 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5u8t | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Structure of Eukaryotic CMG Helicase at a Replication Fork and Implications | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | REPLICATION / CMG helicase / replisome / origin initiation / DNA polymerase / DNA replication | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Unwinding of DNA / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / MCM complex binding / GINS complex / DNA strand elongation involved in mitotic DNA replication / nuclear DNA replication / mitotic DNA replication preinitiation complex assembly / premeiotic DNA replication ...Unwinding of DNA / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / MCM complex binding / GINS complex / DNA strand elongation involved in mitotic DNA replication / nuclear DNA replication / mitotic DNA replication preinitiation complex assembly / premeiotic DNA replication / pre-replicative complex assembly involved in nuclear cell cycle DNA replication / mitotic DNA replication / Activation of the pre-replicative complex / CMG complex / single-stranded 3'-5' DNA helicase activity / nuclear pre-replicative complex / MCM complex / Activation of ATR in response to replication stress / DNA replication preinitiation complex / replication fork protection complex / mitotic DNA replication initiation / double-strand break repair via break-induced replication / silent mating-type cassette heterochromatin formation / regulation of DNA-templated DNA replication initiation / single-stranded DNA helicase activity / DNA strand elongation involved in DNA replication / DNA unwinding involved in DNA replication / nuclear replication fork / 3'-5' DNA helicase activity / DNA replication origin binding / subtelomeric heterochromatin formation / DNA replication initiation / heterochromatin formation / helicase activity / DNA-templated DNA replication / single-stranded DNA binding / DNA helicase / chromosome, telomeric region / DNA damage response / chromatin binding / ATP hydrolysis activity / nucleoplasm / ATP binding / nucleus / metal ion binding / cytoplasm 類似検索 - 分子機能 | |||||||||
| 生物種 |  | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.9 Å | |||||||||
 データ登録者 データ登録者 | Li, B. / Georgescu, R. / Yuan, Z. / Santos, R. / Sun, J. / Zhang, D. / Yurieva, O. / Li, H. / O'Donnell, M.E. | |||||||||
| 資金援助 |  米国, 2件 米国, 2件
| |||||||||
 引用 引用 | ジャーナル: Proc Natl Acad Sci U S A / 年: 2017 タイトル: Structure of eukaryotic CMG helicase at a replication fork and implications to replisome architecture and origin initiation. 著者: Roxana Georgescu / Zuanning Yuan / Lin Bai / Ruda de Luna Almeida Santos / Jingchuan Sun / Dan Zhang / Olga Yurieva / Huilin Li / Michael E O'Donnell / 要旨: The eukaryotic CMG (Cdc45, Mcm2-7, GINS) helicase consists of the Mcm2-7 hexameric ring along with five accessory factors. The Mcm2-7 heterohexamer, like other hexameric helicases, is shaped like a ...The eukaryotic CMG (Cdc45, Mcm2-7, GINS) helicase consists of the Mcm2-7 hexameric ring along with five accessory factors. The Mcm2-7 heterohexamer, like other hexameric helicases, is shaped like a ring with two tiers, an N-tier ring composed of the N-terminal domains, and a C-tier of C-terminal domains; the C-tier contains the motor. In principle, either tier could translocate ahead of the other during movement on DNA. We have used cryo-EM single-particle 3D reconstruction to solve the structure of CMG in complex with a DNA fork. The duplex stem penetrates into the central channel of the N-tier and the unwound leading single-strand DNA traverses the channel through the N-tier into the C-tier motor, 5'-3' through CMG. Therefore, the N-tier ring is pushed ahead by the C-tier ring during CMG translocation, opposite the currently accepted polarity. The polarity of the N-tier ahead of the C-tier places the leading Pol ε below CMG and Pol α-primase at the top of CMG at the replication fork. Surprisingly, the new N-tier to C-tier polarity of translocation reveals an unforeseen quality-control mechanism at the origin. Thus, upon assembly of head-to-head CMGs that encircle double-stranded DNA at the origin, the two CMGs must pass one another to leave the origin and both must remodel onto opposite strands of single-stranded DNA to do so. We propose that head-to-head motors may generate energy that underlies initial melting at the origin. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5u8t.cif.gz | 1 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5u8t.ent.gz | 850.9 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5u8t.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5u8t_validation.pdf.gz | 1.2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5u8t_full_validation.pdf.gz | 1.4 MB | 表示 | |
| XML形式データ | 5u8t_validation.xml.gz | 169.9 KB | 表示 | |
| CIF形式データ | 5u8t_validation.cif.gz | 249.1 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/u8/5u8tftp://data.pdbj.org/pub/pdb/validation_reports/u8/5u8t | HTTPS FTP |
-関連構造データ





-リンク
 PDBj
PDBj













































- 集合体
集合体
| 登録構造単位 | 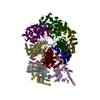
|
|---|---|
| 1 |
|
-要素
-DNA replication licensing factor ... , 5種, 5分子 23467
| #1: タンパク質 | 分子量: 98911.539 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: MCM2, YBL023C, YBL0438 / 発現宿主: |
|---|---|
| #2: タンパク質 | 分子量: 107653.508 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: MCM3, YEL032W, SYGP-ORF23 / 発現宿主: |
| #3: タンパク質 | 分子量: 105138.375 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: MCM4, CDC54, HCD21, YPR019W, YP9531.13 / 発現宿主: |
| #5: タンパク質 | 分子量: 113110.211 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: MCM6, YGL201C / 発現宿主: |
| #6: タンパク質 | 分子量: 95049.875 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: MCM7, CDC47, YBR202W, YBR1441 / 発現宿主: |
-タンパク質 , 2種, 2分子 5E
| #4: タンパク質 | 分子量: 86505.734 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: MCM5, CDC46, YLR274W, L9328.1 / 発現宿主: |
|---|---|
| #11: タンパク質 | 分子量: 74324.836 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: CDC45, SLD4, YLR103C, L8004.11 / 発現宿主: |
-DNA replication complex GINS protein ... , 4種, 4分子 ABCD
| #7: タンパク質 | 分子量: 24230.576 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: PSF1, YDR013W, PZA208, YD8119.18 / 発現宿主: |
|---|---|
| #8: タンパク質 | 分子量: 25096.807 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: PSF2, YJL072C, HRF213, J1086 / 発現宿主: |
| #9: タンパク質 | 分子量: 21977.135 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: PSF3, YOL146W / 発現宿主: |
| #10: タンパク質 | 分子量: 33983.617 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: SLD5, YDR489W / 発現宿主: |
-DNA鎖 / 非ポリマー , 2種, 4分子 F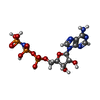
| #12: DNA鎖 | 分子量: 4213.742 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
|---|---|
| #13: 化合物 |  分子量: 506.196 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C10H17N6O12P3 分子量: 506.196 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C10H17N6O12P3コメント: AMP-PNP, エネルギー貯蔵分子類似体*YM |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: CMG-ssDNA / タイプ: COMPLEX / Entity ID: #1-#12 / 由来: RECOMBINANT |
|---|---|
| 分子量 | 値: 0.75 MDa |
| 由来(天然) | 生物種: |
| 由来(組換発現) | 生物種: |
| 緩衝液 | pH: 7.5 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD |
| 撮影 | 電子線照射量: 10 e/Å2 フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: 1.9_1692 / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 4.9 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 395443 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 解像度: 4.9→332.8 Å / SU ML: 1.45 / 位相誤差: 54.63 / 立体化学のターゲット値: MLHL
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å / 溶媒モデル: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル |
|
