- EMDB-8520: Structure of Eukaryotic CMG Helicase at a Replication Fork and Im... -
+
Open data
ID or keywords:
Loading...
-
Basic information
Entry
Database: EMDB / ID: EMD-8520
Title
Structure of Eukaryotic CMG Helicase at a Replication Fork and Implications to Replisome Architecture and Origin Initiation
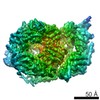
Map data
Eukaryotic CMG Helicase at a Replication Fork
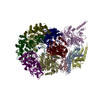
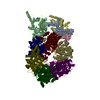
Sample
Complex: CMG-short ssDNA (9 base)
Function / homology
Function and homology information
Unwinding of DNA / DNA strand elongation involved in mitotic DNA replication / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / GINS complex / MCM complex binding / mitotic DNA replication preinitiation complex assembly / nuclear DNA replication / premeiotic DNA replication ...Unwinding of DNA / DNA strand elongation involved in mitotic DNA replication / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / GINS complex / MCM complex binding / mitotic DNA replication preinitiation complex assembly / nuclear DNA replication / premeiotic DNA replication / pre-replicative complex assembly involved in nuclear cell cycle DNA replication / Activation of the pre-replicative complex / mitotic DNA replication / nuclear pre-replicative complex / CMG complex / DNA replication preinitiation complex / Activation of ATR in response to replication stress / double-strand break repair via break-induced replication / MCM complex / mitotic DNA replication initiation / silent mating-type cassette heterochromatin formation / single-stranded DNA helicase activity / regulation of DNA-templated DNA replication initiation / DNA strand elongation involved in DNA replication / nuclear replication fork / DNA replication origin binding / DNA replication initiation / subtelomeric heterochromatin formation / DNA helicase activity / helicase activity / transcription elongation by RNA polymerase II / DNA-templated DNA replication / peroxisome / heterochromatin formation / single-stranded DNA binding / DNA helicase / DNA replication / chromosome, telomeric region / chromatin binding / DNA damage response / ATP hydrolysis activity / zinc ion binding / nucleoplasm / ATP binding / nucleus / cytoplasm Similarity search - Function
: / PSF2 N-terminal domain / : / : / PSF3 N-terminal domain / PSF1 C-terminal domain / DNA replication complex GINS protein Psf2 / CDC45 family / CDC45 / GINS complex, subunit Psf3 ...: / PSF2 N-terminal domain / : / : / PSF3 N-terminal domain / PSF1 C-terminal domain / DNA replication complex GINS protein Psf2 / CDC45 family / CDC45 / GINS complex, subunit Psf3 / DNA replication complex GINS protein SLD5, C-terminal / GINS complex, subunit Psf3 superfamily / GINS complex protein Sld5, alpha-helical domain / DNA replication complex GINS protein SLD5 C-terminus / GINS complex subunit Sld5 / GINS subunit, domain A / GINS complex protein helical bundle domain / GINS complex, subunit Psf1 / : / MCM3 winged helix domain / GINS, helical bundle-like domain superfamily / : / MCM5, C-terminal domain / DNA replication licensing factor MCM7, winged helix / DNA replication licensing factor Mcm5 / MCM4, winged helix domain / DNA replication licensing factor Mcm3 / Mini-chromosome maintenance complex protein 4 / : / MCM3-like, winged helix domain / DNA replication licensing factor Mcm6 / DNA replication licensing factor Mcm7 / Mcm6, C-terminal winged-helix domain / MCM6 C-terminal winged-helix domain / DNA replication licensing factor Mcm2 / Mini-chromosome maintenance protein 2 / Mini-chromosome maintenance, conserved site / MCM family signature. / MCM N-terminal domain / MCM N-terminal domain / MCM OB domain / MCM OB domain / Mini-chromosome maintenance protein / MCM, AAA-lid domain / MCM P-loop domain / MCM AAA-lid domain / MCM family C-terminal AAA(+) ATPase domain (MCM-CTD) profile. / minichromosome maintenance proteins / MCM domain / Winged helix-like DNA-binding domain superfamily / Nucleic acid-binding, OB-fold / ATPases associated with a variety of cellular activities / AAA+ ATPase domain / P-loop containing nucleoside triphosphate hydrolase Similarity search - Domain/homology
DNA replication licensing factor MCM3 / DNA replication licensing factor MCM2 / Minichromosome maintenance protein 5 / DNA replication licensing factor MCM4 / DNA replication licensing factor MCM7 / DNA replication complex GINS protein PSF2 / DNA replication licensing factor MCM6 / DNA replication complex GINS protein SLD5 / Cell division control protein 45 / DNA replication complex GINS protein PSF3 / DNA replication complex GINS protein PSF1 Similarity search - Component
Biological species
Saccharomyces cerevisiae (brewer's yeast)
Method
single particle reconstruction / cryo EM / Resolution: 6.2 Å
National Institutes of Health/National Institute of General Medical Sciences (NIH/NIGMS)
GM111472
United States
National Institutes of Health/National Institute of General Medical Sciences (NIH/NIGMS)
GM115809
United States
Citation
Journal: Proc Natl Acad Sci U S A / Year: 2017 Title: Structure of eukaryotic CMG helicase at a replication fork and implications to replisome architecture and origin initiation. Authors: Roxana Georgescu / Zuanning Yuan / Lin Bai / Ruda de Luna Almeida Santos / Jingchuan Sun / Dan Zhang / Olga Yurieva / Huilin Li / Michael E O'Donnell / Abstract: The eukaryotic CMG (Cdc45, Mcm2-7, GINS) helicase consists of the Mcm2-7 hexameric ring along with five accessory factors. The Mcm2-7 heterohexamer, like other hexameric helicases, is shaped like a ...The eukaryotic CMG (Cdc45, Mcm2-7, GINS) helicase consists of the Mcm2-7 hexameric ring along with five accessory factors. The Mcm2-7 heterohexamer, like other hexameric helicases, is shaped like a ring with two tiers, an N-tier ring composed of the N-terminal domains, and a C-tier of C-terminal domains; the C-tier contains the motor. In principle, either tier could translocate ahead of the other during movement on DNA. We have used cryo-EM single-particle 3D reconstruction to solve the structure of CMG in complex with a DNA fork. The duplex stem penetrates into the central channel of the N-tier and the unwound leading single-strand DNA traverses the channel through the N-tier into the C-tier motor, 5'-3' through CMG. Therefore, the N-tier ring is pushed ahead by the C-tier ring during CMG translocation, opposite the currently accepted polarity. The polarity of the N-tier ahead of the C-tier places the leading Pol ε below CMG and Pol α-primase at the top of CMG at the replication fork. Surprisingly, the new N-tier to C-tier polarity of translocation reveals an unforeseen quality-control mechanism at the origin. Thus, upon assembly of head-to-head CMGs that encircle double-stranded DNA at the origin, the two CMGs must pass one another to leave the origin and both must remodel onto opposite strands of single-stranded DNA to do so. We propose that head-to-head motors may generate energy that underlies initial melting at the origin.
History
Deposition
Dec 15, 2016
-
Header (metadata) release
Jan 11, 2017
-
Map release
Jan 25, 2017
-
Update
Jan 29, 2020
-
Current status
Jan 29, 2020
Processing site: RCSB / Status: Released
-
Structure visualization
Movie
Surface view with section colored by density value
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller

 Yorodumi
Yorodumi Open data
Open data
 Basic information
Basic information Map data
Map data Sample
Sample Function and homology information
Function and homology information
 Authors
Authors United States, 2 items
United States, 2 items  Citation
Citation Structure visualization
Structure visualization
 Downloads & links
Downloads & links emd_8520.png
emd_8520.png http://ftp.pdbj.org/pub/emdb/structures/EMD-8520
http://ftp.pdbj.org/pub/emdb/structures/EMD-8520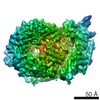

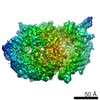
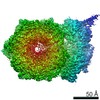
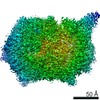


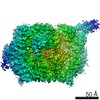


 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















 Sample components
Sample components Processing
Processing Electron microscopy
Electron microscopy FIELD EMISSION GUN
FIELD EMISSION GUN
