 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-5ty4: MicroED structure of a complex between monomeric TGF-b and its re... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5ty4 | ||||||
|---|---|---|---|---|---|---|---|
| Title | MicroED structure of a complex between monomeric TGF-b and its receptor, TbRII, at 2.9 A resolution | ||||||
 Components Components |
| ||||||
 Keywords Keywords | TRANSFERASE | ||||||
| Function / homology |  Function and homology information Function and homology informationregulation of timing of catagen / positive regulation of activation-induced cell death of T cells / regulation of apoptotic process involved in outflow tract morphogenesis / negative regulation of epithelial to mesenchymal transition involved in endocardial cushion formation / ascending aorta morphogenesis / cardioblast differentiation / uterine wall breakdown / positive regulation of tolerance induction to self antigen / positive regulation of B cell tolerance induction / bronchus morphogenesis ...regulation of timing of catagen / positive regulation of activation-induced cell death of T cells / regulation of apoptotic process involved in outflow tract morphogenesis / negative regulation of epithelial to mesenchymal transition involved in endocardial cushion formation / ascending aorta morphogenesis / cardioblast differentiation / uterine wall breakdown / positive regulation of tolerance induction to self antigen / positive regulation of B cell tolerance induction / bronchus morphogenesis / inferior endocardial cushion morphogenesis / transforming growth factor beta receptor activity, type II / positive regulation of timing of catagen / mammary gland morphogenesis / lens fiber cell apoptotic process / growth plate cartilage chondrocyte growth / positive regulation of cardioblast differentiation / substantia propria of cornea development / tricuspid valve morphogenesis / TGFBR2 MSI Frameshift Mutants in Cancer / cardiac right ventricle morphogenesis / miRNA transport / regulation of transforming growth factor beta2 production / atrial septum morphogenesis / transforming growth factor beta ligand-receptor complex / type III transforming growth factor beta receptor binding / pharyngeal arch artery morphogenesis / positive regulation of epithelial to mesenchymal transition involved in endocardial cushion formation / aorta morphogenesis / Langerhans cell differentiation / positive regulation of heart contraction / transforming growth factor beta receptor activity / TGFBR2 Kinase Domain Mutants in Cancer / glial cell migration / cardiac left ventricle morphogenesis / negative regulation of macrophage cytokine production / positive regulation of extracellular matrix disassembly / activation-induced cell death of T cells / secondary palate development / SMAD2/3 Phosphorylation Motif Mutants in Cancer / TGFBR1 KD Mutants in Cancer / positive regulation of integrin biosynthetic process / endocardial cushion fusion / atrial septum primum morphogenesis / positive regulation of T cell tolerance induction / negative regulation of cartilage development / positive regulation of NK T cell differentiation / cardiac epithelial to mesenchymal transition / somatic stem cell division / membranous septum morphogenesis / heart valve morphogenesis / signaling / positive regulation of stress-activated MAPK cascade / pericyte cell differentiation / TGFBR3 regulates TGF-beta signaling / neuron fate commitment / activin receptor activity, type I / activin receptor complex / regulation of stem cell proliferation / lung lobe morphogenesis / embryonic digestive tract development / type II transforming growth factor beta receptor binding / eye development / receptor protein serine/threonine kinase / activin binding / neural retina development / cranial skeletal system development / pulmonary valve morphogenesis / transmembrane receptor protein serine/threonine kinase activity / transforming growth factor beta receptor binding / TGFBR1 LBD Mutants in Cancer / SMAD protein signal transduction / embryonic cranial skeleton morphogenesis / myeloid dendritic cell differentiation / ventricular trabecula myocardium morphogenesis / glycosaminoglycan binding / activin receptor signaling pathway / type I transforming growth factor beta receptor binding / positive regulation of CD4-positive, alpha-beta T cell proliferation / negative regulation of Ras protein signal transduction / positive regulation of extrinsic apoptotic signaling pathway in absence of ligand / outflow tract septum morphogenesis / regulation of stem cell differentiation / embryo development ending in birth or egg hatching / response to cholesterol / cell-cell junction organization / transforming growth factor beta binding / collagen fibril organization / lens development in camera-type eye / positive regulation of cell adhesion mediated by integrin / atrioventricular valve morphogenesis / aortic valve morphogenesis / kinase activator activity / embryonic limb morphogenesis / face morphogenesis / artery morphogenesis / embryonic hemopoiesis / odontogenesis / positive regulation of mesenchymal cell proliferation / endocardial cushion morphogenesis Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method | ELECTRON CRYSTALLOGRAPHY / electron crystallography /  MOLECULAR REPLACEMENT / cryo EM / Resolution: 2.9 Å MOLECULAR REPLACEMENT / cryo EM / Resolution: 2.9 Å | ||||||
 Authors Authors | Weiss, S.C. / de la Cruz, M.J. / Hattne, J. / Shi, D. / Reyes, F.E. / Callero, G. / Gonen, T. | ||||||
 Citation Citation | Journal: Nat Methods / Year: 2017 Title: Atomic-resolution structures from fragmented protein crystals with the cryoEM method MicroED. Authors: M Jason de la Cruz / Johan Hattne / Dan Shi / Paul Seidler / Jose Rodriguez / Francis E Reyes / Michael R Sawaya / Duilio Cascio / Simon C Weiss / Sun Kyung Kim / Cynthia S Hinck / Andrew P ...Authors: M Jason de la Cruz / Johan Hattne / Dan Shi / Paul Seidler / Jose Rodriguez / Francis E Reyes / Michael R Sawaya / Duilio Cascio / Simon C Weiss / Sun Kyung Kim / Cynthia S Hinck / Andrew P Hinck / Guillermo Calero / David Eisenberg / Tamir Gonen /  Abstract: Traditionally, crystallographic analysis of macromolecules has depended on large, well-ordered crystals, which often require significant effort to obtain. Even sizable crystals sometimes suffer from ...Traditionally, crystallographic analysis of macromolecules has depended on large, well-ordered crystals, which often require significant effort to obtain. Even sizable crystals sometimes suffer from pathologies that render them inappropriate for high-resolution structure determination. Here we show that fragmentation of large, imperfect crystals into microcrystals or nanocrystals can provide a simple path for high-resolution structure determination by the cryoEM method MicroED and potentially by serial femtosecond crystallography. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5ty4.cif.gz | 51.1 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5ty4.ent.gz | 32.6 KB | Display | PDB format |
| PDBx/mmJSON format | 5ty4.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ty/5ty4ftp://data.pdbj.org/pub/pdb/validation_reports/ty/5ty4 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  8472MC  8216C  8217C  8218C  8219C  8220C  8221C  8222C  5k7nC  5k7oC  5k7pC  5k7qC  5k7rC  5k7sC  5k7tC  1ktzS |
|---|---|
| Similar structure data | |
| Experimental dataset #1 | Data reference: 10.15785/SBGRID/368 / Data set type: diffraction image data |








-Links
 PDBj
PDBj








- Assembly
Assembly
| Deposited unit | 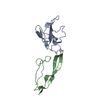
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 11788.519 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: TGFBR2 / Production host:  References: UniProt: P37173, receptor protein serine/threonine kinase |
|---|---|
| #2: Protein | Mass: 11076.813 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Production host: |
| Has protein modification | Y |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON CRYSTALLOGRAPHY / Number of used crystals: 1 |
|---|---|
| EM experiment | Aggregation state: 3D ARRAY / 3D reconstruction method: electron crystallography |
- Sample preparation
Sample preparation
| Component | Name: Complex between monomeric TGF-b and its receptor, TbRII Type: COMPLEX / Entity ID: all / Source: NATURAL |
|---|---|
| Molecular weight | Value: 0.019072 MDa / Experimental value: NO |
| Source (natural) | Organism: Homo sapiens (human) |
| Buffer solution | pH: 7.5 |
| Buffer component | Conc.: 100 mM / Name: HEPES |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Cryogen name: ETHANE |
| Crystal | Density Matthews: 2.58 Å3/Da / Density % sol: 52.24 % |
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, hanging drop / pH: 7.5 Details: 0.5 ul 20 mg/mL protein + 0.25 ul mother liquor + 0.2 ul seed stock in 100 mM HEPES/NaOH pH 7.5, 45% MPD |
-Data collection
| Experimental equipment |  Model: Tecnai F20 / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TECNAI F20 |
| Electron gun | Electron source: FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: DIFFRACTION |
| Specimen holder | Cryogen: NITROGEN |
| Image recording | Average exposure time: 4.1 sec. / Electron dose: 0.004 e/Å2 / Film or detector model: TVIPS TEMCAM-F416 (4k x 4k) / Num. of diffraction images: 353 / Num. of grids imaged: 2 / Num. of real images: 353 |
| Image scans | Sampling size: 0.0311999992 µm / Width: 2048 / Height: 2048 |
| EM diffraction | Camera length: 2000 mm |
| EM diffraction shell | Resolution: 2.9→3.65 Å / Fourier space coverage: 69.1 % / Multiplicity: 3.9 / Num. of structure factors: 1884 / Phase residual: 46.4 ° |
| EM diffraction stats | Fourier space coverage: 71.9 % / High resolution: 2.9 Å / Num. of intensities measured: 14911 / Num. of structure factors: 3884 / Phase error: 30.99 ° / Phase residual: 43.53 ° / Phase error rejection criteria: 0 / Rmerge: 0.293 / Rsym: 0.293 |
| Diffraction | Mean temperature: 293 K |
| Diffraction source | Source: ELECTRON MICROSCOPE / Type: OTHER / Wavelength: 0.0250793397 Å |
| Detector | Type: TVIPS TEMCAM-F416 / Detector: CMOS / Date: May 4, 2016 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: electron |
| Radiation wavelength | Wavelength: 0.0250793397 Å / Relative weight: 1 |
| Reflection | Resolution: 2.9→26.64 Å / Num. obs: 3884 / % possible obs: 71.9 % / Redundancy: 3.8 % / Biso Wilson estimate: 64 Å2 / CC1/2: 0.951 / Rmerge(I) obs: 0.293 / Rsym value: 0.293 / Net I/σ(I): 3.3 |
| Reflection shell | Resolution: 2.9→3.07 Å / Redundancy: 3.9 % / Rmerge(I) obs: 2.024 / Mean I/σ(I) obs: 0.8 / CC1/2: 0.255 / % possible all: 71.3 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| |||||||||||||||||||||||||||||||||||||||||||||
| EM 3D crystal entity | ∠α: 90 ° / ∠β: 90 ° / ∠γ: 90 ° / A: 41.5298 Å / B: 71.3297 Å / C: 79.5082 Å / Space group name: P212121 / Space group num: 19 | |||||||||||||||||||||||||||||||||||||||||||||
| CTF correction | Type: NONE | |||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 2.9 Å / Resolution method: DIFFRACTION PATTERN/LAYERLINES / Symmetry type: 3D CRYSTAL | |||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: OTHER / Space: RECIPROCAL | |||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: PDB entry 1KTZ Resolution: 2.9→26.64 Å / SU ML: 0.4 / Cross valid method: FREE R-VALUE / Phase error: 30.99
| |||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å | |||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.9→26.64 Å
| |||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
