UBC13-MMS2 complex / ubiquitin conjugating enzyme complex / ubiquitin-protein transferase activator activity / sperm DNA condensation / positive regulation of protein K63-linked ubiquitination / DNA double-strand break processing / protein K6-linked ubiquitination / isotype switching / hypothalamus gonadotrophin-releasing hormone neuron development / DNA damage tolerance ...UBC13-MMS2 complex / ubiquitin conjugating enzyme complex / ubiquitin-protein transferase activator activity / sperm DNA condensation / positive regulation of protein K63-linked ubiquitination / DNA double-strand break processing / protein K6-linked ubiquitination / isotype switching / hypothalamus gonadotrophin-releasing hormone neuron development / DNA damage tolerance / female meiosis I / positive regulation of protein monoubiquitination / fat pad development / mitochondrion transport along microtubule / E2 ubiquitin-conjugating enzyme / response to ionizing radiation / female gonad development / positive regulation of double-strand break repair / DNA repair-dependent chromatin remodeling / seminiferous tubule development / male meiosis I / ubiquitin conjugating enzyme activity / positive regulation of intracellular signal transduction / negative regulation of transcription elongation by RNA polymerase II / positive regulation of intrinsic apoptotic signaling pathway by p53 class mediator / signal transduction in response to DNA damage / protein K63-linked ubiquitination / protein monoubiquitination / ubiquitin ligase complex / interstrand cross-link repair / protein K48-linked ubiquitination / regulation of DNA repair / positive regulation of double-strand break repair via homologous recombination / protein autoubiquitination / energy homeostasis / regulation of neuron apoptotic process / neuron projection morphogenesis / negative regulation of TORC1 signaling / regulation of proteasomal protein catabolic process / Maturation of protein E / Maturation of protein E / ER Quality Control Compartment (ERQC) / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / Prevention of phagosomal-lysosomal fusion / IRAK2 mediated activation of TAK1 complex / Alpha-protein kinase 1 signaling pathway / Glycogen synthesis / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / antiviral innate immune response / Endosomal Sorting Complex Required For Transport (ESCRT) / Membrane binding and targetting of GAG proteins / Negative regulation of FLT3 / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Regulation of TBK1, IKKε-mediated activation of IRF3, IRF7 upon TLR3 ligation / Constitutive Signaling by NOTCH1 HD Domain Mutants / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1,TRAF6-dependent induction of TAK1 complex / TICAM1-dependent activation of IRF3/IRF7 / APC/C:Cdc20 mediated degradation of Cyclin B / Downregulation of ERBB4 signaling / Regulation of FZD by ubiquitination / APC-Cdc20 mediated degradation of Nek2A / p75NTR recruits signalling complexes / InlA-mediated entry of Listeria monocytogenes into host cells / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / TRAF6-mediated induction of TAK1 complex within TLR4 complex / Regulation of pyruvate metabolism / NF-kB is activated and signals survival / Regulation of innate immune responses to cytosolic DNA / Downregulation of ERBB2:ERBB3 signaling / Pexophagy / NRIF signals cell death from the nucleus / VLDLR internalisation and degradation / Regulation of PTEN localization / positive regulation of DNA repair / Activated NOTCH1 Transmits Signal to the Nucleus / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Regulation of BACH1 activity / MAP3K8 (TPL2)-dependent MAPK1/3 activation / TICAM1, RIP1-mediated IKK complex recruitment / Translesion synthesis by REV1 / InlB-mediated entry of Listeria monocytogenes into host cell / Translesion synthesis by POLK / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / Downregulation of TGF-beta receptor signaling / epigenetic regulation of gene expression / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / Josephin domain DUBs / Translesion synthesis by POLI / IKK complex recruitment mediated by RIP1 / Regulation of activated PAK-2p34 by proteasome mediated degradation / Gap-filling DNA repair synthesis and ligation in GG-NER / ubiquitin binding / positive regulation of protein ubiquitination / PINK1-PRKN Mediated Mitophagy / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) 類似検索 - 分子機能
ジャーナル: J Biol Chem / 年: 2016 タイトル: RNF8 E3 Ubiquitin Ligase Stimulates Ubc13 E2 Conjugating Activity That Is Essential for DNA Double Strand Break Signaling and BRCA1 Tumor Suppressor Recruitment. 著者: Curtis D Hodge / Ismail H Ismail / Ross A Edwards / Greg L Hura / Andrew T Xiao / John A Tainer / Michael J Hendzel / J N Mark Glover / 要旨: DNA double strand break (DSB) responses depend on the sequential actions of the E3 ubiquitin ligases RNF8 and RNF168 plus E2 ubiquitin-conjugating enzyme Ubc13 to specifically generate histone Lys-63- ...DNA double strand break (DSB) responses depend on the sequential actions of the E3 ubiquitin ligases RNF8 and RNF168 plus E2 ubiquitin-conjugating enzyme Ubc13 to specifically generate histone Lys-63-linked ubiquitin chains in DSB signaling. Here, we defined the activated RNF8-Ubc13∼ubiquitin complex by x-ray crystallography and its functional solution conformations by x-ray scattering, as tested by separation-of-function mutations imaged in cells by immunofluorescence. The collective results show that the RING E3 RNF8 targets E2 Ubc13 to DSB sites and plays a critical role in damage signaling by stimulating polyubiquitination through modulating conformations of ubiquitin covalently linked to the Ubc13 active site. Structure-guided separation-of-function mutations show that the RNF8 E2 stimulating activity is essential for DSB signaling in mammalian cells and is necessary for downstream recruitment of 53BP1 and BRCA1. Chromatin-targeted RNF168 rescues 53BP1 recruitment involved in non-homologous end joining but not BRCA1 recruitment for homologous recombination. These findings suggest an allosteric approach to targeting the ubiquitin-docking cleft at the E2-E3 interface for possible interventions in cancer and chronic inflammation, and moreover, they establish an independent RNF8 role in BRCA1 recruitment.
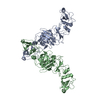
The biological unit is a dimer set (2RNF8 protomers, 2Ubc13~Ub). There are 2 biological units in the asymmetric unit (chains A, B, C, D, E, F and chains G, H, I, J, K, L
-
要素
#1: タンパク質
Ubiquitin-conjugatingenzymeE2N / Bendless-like ubiquitin-conjugating enzyme / Ubc13 / UbcH13 / Ubiquitin carrier protein N / ...Bendless-like ubiquitin-conjugating enzyme / Ubc13 / UbcH13 / Ubiquitin carrier protein N / Ubiquitin-protein ligase N
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) X線回折 /
X線回折 /  データ登録者
データ登録者 引用
引用


 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード






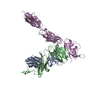

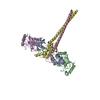
 PDBj
PDBj



















 集合体
集合体
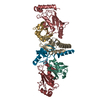


 分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn
分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn 試料調製
試料調製 解析
解析