 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
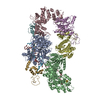
Yorodumi- PDB-4o02: AlphaVBeta3 integrin in complex with monoclonal antibody FAB fragment. -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 4o02 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | AlphaVBeta3 integrin in complex with monoclonal antibody FAB fragment. | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | PROTEIN BINDING | |||||||||
| Function / homology |  Function and homology information Function and homology informationintegrin alphav-beta8 complex / integrin alphav-beta6 complex / transforming growth factor beta production / negative regulation of entry of bacterium into host cell / integrin alphav-beta5 complex / opsonin binding / integrin alphav-beta1 complex / Cross-presentation of particulate exogenous antigens (phagosomes) / extracellular matrix protein binding / regulation of serotonin uptake ...integrin alphav-beta8 complex / integrin alphav-beta6 complex / transforming growth factor beta production / negative regulation of entry of bacterium into host cell / integrin alphav-beta5 complex / opsonin binding / integrin alphav-beta1 complex / Cross-presentation of particulate exogenous antigens (phagosomes) / extracellular matrix protein binding / regulation of serotonin uptake / positive regulation of adenylate cyclase-inhibiting opioid receptor signaling pathway / tube development / alpha9-beta1 integrin-ADAM8 complex / regulation of trophoblast cell migration / integrin alphaIIb-beta3 complex / regulation of postsynaptic neurotransmitter receptor diffusion trapping / maintenance of postsynaptic specialization structure / alphav-beta3 integrin-vitronectin complex / regulation of extracellular matrix organization / platelet alpha granule membrane / Laminin interactions / positive regulation of glomerular mesangial cell proliferation / integrin alphav-beta3 complex / negative regulation of lipoprotein metabolic process / alphav-beta3 integrin-PKCalpha complex / positive regulation of small GTPase mediated signal transduction / fibrinogen binding / entry into host cell by a symbiont-containing vacuole / alphav-beta3 integrin-HMGB1 complex / negative regulation of lipid transport / vascular endothelial growth factor receptor 2 binding / regulation of phagocytosis / angiogenesis involved in wound healing / positive regulation of vascular endothelial growth factor signaling pathway / cell-substrate junction assembly / regulation of release of sequestered calcium ion into cytosol / positive regulation of bone resorption / mesodermal cell differentiation / Elastic fibre formation / alphav-beta3 integrin-IGF-1-IGF1R complex / transforming growth factor beta binding / platelet-derived growth factor receptor binding / filopodium membrane / glycinergic synapse / extracellular matrix binding / positive regulation of fibroblast migration / positive regulation of cell adhesion mediated by integrin / positive regulation of vascular endothelial growth factor receptor signaling pathway / apolipoprotein A-I-mediated signaling pathway / smooth muscle cell migration / regulation of bone resorption / negative regulation of low-density lipoprotein particle clearance / apoptotic cell clearance / wound healing, spreading of epidermal cells / positive regulation of smooth muscle cell migration / integrin complex / Molecules associated with elastic fibres / heterotypic cell-cell adhesion / cell adhesion mediated by integrin / endodermal cell differentiation / negative chemotaxis / positive regulation of osteoblast proliferation / platelet-derived growth factor receptor signaling pathway / Mechanical load activates signaling by PIEZO1 and integrins in osteocytes / positive regulation of cell-matrix adhesion / Syndecan interactions / p130Cas linkage to MAPK signaling for integrins / cellular response to insulin-like growth factor stimulus / regulation of postsynaptic neurotransmitter receptor internalization / cell-substrate adhesion / protein disulfide isomerase activity / microvillus membrane / PECAM1 interactions / GRB2:SOS provides linkage to MAPK signaling for Integrins / TGF-beta receptor signaling activates SMADs / lamellipodium membrane / fibronectin binding / positive regulation of intracellular signal transduction / negative regulation of macrophage derived foam cell differentiation / negative regulation of lipid storage / vasculogenesis / blood coagulation, fibrin clot formation / negative regulation of endothelial cell apoptotic process / extrinsic apoptotic signaling pathway in absence of ligand / ERK1 and ERK2 cascade / ECM proteoglycans / Integrin cell surface interactions / positive regulation of T cell migration / cellular response to platelet-derived growth factor stimulus / embryo implantation / specific granule membrane / coreceptor activity / substrate adhesion-dependent cell spreading / Integrin signaling / positive regulation of substrate adhesion-dependent cell spreading / phagocytic vesicle / positive regulation of endothelial cell proliferation / cell-matrix adhesion / positive regulation of smooth muscle cell proliferation / cell adhesion molecule binding Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.605 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.605 Å | |||||||||
 Authors Authors | Mahalingam, B. / van Agthoven, J. / Xiong, J. / Arnaout, M.A. | |||||||||
 Citation Citation | Journal: J.Biol.Chem. / Year: 2014 Title: Atomic basis for the species-specific inhibition of alpha V integrins by monoclonal antibody 17E6 is revealed by the crystal structure of alpha V beta 3 ectodomain-17E6 Fab complex. Authors: Mahalingam, B. / Van Agthoven, J.F. / Xiong, J.P. / Alonso, J.L. / Adair, B.D. / Rui, X. / Anand, S. / Mehrbod, M. / Mofrad, M.R. / Burger, C. / Goodman, S.L. / Arnaout, M.A. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 4o02.cif.gz | 731.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb4o02.ent.gz | 590.3 KB | Display | PDB format |
| PDBx/mmJSON format | 4o02.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/o0/4o02ftp://data.pdbj.org/pub/pdb/validation_reports/o0/4o02 | HTTPS FTP |
|---|
-Related structure data
| Similar structure data |
|---|



-Links
 PDBj
PDBj











- Assembly
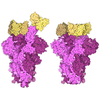
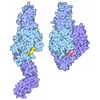
Assembly
| Deposited unit | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
|
-Components
-Protein , 2 types, 2 molecules AB
| #1: Protein | Mass: 106375.805 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ITGAV, MSK8, VNRA / Production host:   Spodoptera frugiperda (fall armyworm) / References: UniProt: P06756 Spodoptera frugiperda (fall armyworm) / References: UniProt: P06756 |
|---|---|
| #2: Protein | Mass: 76523.125 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ITGB3, GP3A / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: P05106 |
-Antibody , 2 types, 2 molecules LH
| #3: Antibody | Mass: 23706.059 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #4: Antibody | Mass: 23298.984 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Sugars , 6 types, 10 molecules 
| #5: Polysaccharide | beta-D-mannopyranose-(1-4)-alpha-D-mannopyranose-(1-6)-[alpha-D-mannopyranose-(1-3)]beta-D- ...beta-D-mannopyranose-(1-4)-alpha-D-mannopyranose-(1-6)-[alpha-D-mannopyranose-(1-3)]beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose Source method: isolated from a genetically manipulated source | ||||
|---|---|---|---|---|---|
| #6: Polysaccharide | beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta- ...beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose Source method: isolated from a genetically manipulated source | ||||
| #7: Polysaccharide | alpha-D-mannopyranose-(1-3)-alpha-D-mannopyranose-(1-6)-[alpha-D-mannopyranose-(1-3)]beta-D- ...alpha-D-mannopyranose-(1-3)-alpha-D-mannopyranose-(1-6)-[alpha-D-mannopyranose-(1-3)]beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose Source method: isolated from a genetically manipulated source | ||||
| #8: Polysaccharide | Source method: isolated from a genetically manipulated source #9: Polysaccharide | alpha-D-mannopyranose-(1-3)-alpha-D-mannopyranose-(1-3)-alpha-D-mannopyranose-(1-3)-beta-D- ...alpha-D-mannopyranose-(1-3)-alpha-D-mannopyranose-(1-3)-alpha-D-mannopyranose-(1-3)-beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose | Source method: isolated from a genetically manipulated source #10: Sugar |  Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 3 Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 3Source method: isolated from a genetically manipulated source Formula: C8H15NO6 |
-Non-polymers , 2 types, 7 molecules 
| #11: Chemical | ChemComp-MN /  Mass: 54.938 Da / Num. of mol.: 5 / Source method: obtained synthetically / Formula: Mn Mass: 54.938 Da / Num. of mol.: 5 / Source method: obtained synthetically / Formula: Mn#12: Chemical |  Mass: 62.005 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: NO3 Mass: 62.005 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: NO3 |
|---|
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 3.28 Å3/Da / Density % sol: 62.46 % |
|---|---|
| Crystal grow | Temperature: 298 K / Method: vapor diffusion, hanging drop / pH: 5.6 Details: 13.5% PEG 4000, pH 5.6, VAPOR DIFFUSION, HANGING DROP, temperature 298K |
-Data collection
| Diffraction source | Source: SYNCHROTRON / Site: APS  / Beamline: 19-ID / Wavelength: 0.97857 / Beamline: 19-ID / Wavelength: 0.97857 |
|---|---|
| Detector | Type: ADSC QUANTUM 315r / Detector: CCD |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.97857 Å / Relative weight: 1 |
| Reflection | Resolution: 3.6→50 Å / Num. obs: 31351 / % possible obs: 87.9 % / Observed criterion σ(I): 31351 / Redundancy: 3.8 % / Biso Wilson estimate: 112 Å2 / Rsym value: 0.131 / Net I/σ(I): 8.3 |
| Reflection shell | Resolution: 3.6→3.73 Å / Redundancy: 2.8 % / Mean I/σ(I) obs: 1 / Rsym value: 0.866 / % possible all: 55.2 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT / Resolution: 3.605→49.862 Å / SU ML: 0.53 / σ(F): 1.34 / Phase error: 33.92 / Stereochemistry target values: ML / Details: RAMACHANDRAN RESTRAINS
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 3.605→49.862 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|