 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-4g3x: Crystal Structure of Q61L H-Ras-GppNHp bound to the RBD of Raf Kinase -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 4g3x | ||||||
|---|---|---|---|---|---|---|---|
| Title | Crystal Structure of Q61L H-Ras-GppNHp bound to the RBD of Raf Kinase | ||||||
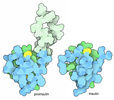
 Components Components |
| ||||||
 Keywords Keywords | HYDROLASE/TRANSFERASE / H-Ras / Ras / Raf kinase / Raf / GTPase / allosteric regulation / intrinsic hydrolysis / protein-protein interaction / kinase / GTP binding / HYDROLASE-TRANSFERASE complex | ||||||
| Function / homology |  Function and homology information Function and homology informationGTPase complex / phospholipase C activator activity / regulation of Rho protein signal transduction / SHOC2 M1731 mutant abolishes MRAS complex function / Gain-of-function MRAS complexes activate RAF signaling / Rap1 signalling / oncogene-induced cell senescence / positive regulation of miRNA metabolic process / positive regulation of ruffle assembly / T-helper 1 type immune response ...GTPase complex / phospholipase C activator activity / regulation of Rho protein signal transduction / SHOC2 M1731 mutant abolishes MRAS complex function / Gain-of-function MRAS complexes activate RAF signaling / Rap1 signalling / oncogene-induced cell senescence / positive regulation of miRNA metabolic process / positive regulation of ruffle assembly / T-helper 1 type immune response / Negative feedback regulation of MAPK pathway / defense response to protozoan / IFNG signaling activates MAPKs / GP1b-IX-V activation signalling / positive regulation of wound healing / ERBB2-ERBB3 signaling pathway / adipose tissue development / regulation of neurotransmitter receptor localization to postsynaptic specialization membrane / neuromuscular junction development / regulation of cell differentiation / pseudopodium / Signaling by RAS GAP mutants / Signaling by RAS GTPase mutants / Activation of RAS in B cells / RAS signaling downstream of NF1 loss-of-function variants / positive regulation of peptidyl-serine phosphorylation / MAP kinase kinase kinase activity / positive regulation of protein targeting to membrane / SOS-mediated signalling / Activated NTRK3 signals through RAS / Activated NTRK2 signals through RAS / type II interferon-mediated signaling pathway / SHC1 events in ERBB4 signaling / Schwann cell development / Signalling to RAS / negative regulation of protein-containing complex assembly / SHC-related events triggered by IGF1R / Activated NTRK2 signals through FRS2 and FRS3 / Estrogen-stimulated signaling through PRKCZ / SHC-mediated cascade:FGFR3 / MET activates RAS signaling / SHC-mediated cascade:FGFR2 / SHC-mediated cascade:FGFR4 / Signaling by PDGFRA transmembrane, juxtamembrane and kinase domain mutants / Signaling by PDGFRA extracellular domain mutants / PTK6 Regulates RHO GTPases, RAS GTPase and MAP kinases / Erythropoietin activates RAS / SHC-mediated cascade:FGFR1 / myelination / Signaling by FGFR4 in disease / fibroblast proliferation / FRS-mediated FGFR3 signaling / Signaling by FLT3 ITD and TKD mutants / FRS-mediated FGFR2 signaling / FRS-mediated FGFR4 signaling / p38MAPK events / Signaling by FGFR3 in disease / FRS-mediated FGFR1 signaling / Tie2 Signaling / insulin-like growth factor receptor signaling pathway / protein-membrane adaptor activity / Signaling by FGFR2 in disease / EPHB-mediated forward signaling / Signaling by FLT3 fusion proteins / GRB2 events in EGFR signaling / SHC1 events in EGFR signaling / FLT3 Signaling / animal organ morphogenesis / Signaling by FGFR1 in disease / EGFR Transactivation by Gastrin / NCAM signaling for neurite out-growth / CD209 (DC-SIGN) signaling / intrinsic apoptotic signaling pathway / positive regulation of epithelial cell proliferation / Downstream signal transduction / GRB2 events in ERBB2 signaling / Insulin receptor signalling cascade / SHC1 events in ERBB2 signaling / Constitutive Signaling by Overexpressed ERBB2 / Ras activation upon Ca2+ influx through NMDA receptor / Signaling by phosphorylated juxtamembrane, extracellular and kinase domain KIT mutants / adenylate cyclase activator activity / VEGFR2 mediated cell proliferation / wound healing / regulation of actin cytoskeleton organization / small monomeric GTPase / FCERI mediated MAPK activation / regulation of long-term neuronal synaptic plasticity / Signaling by ERBB2 TMD/JMD mutants / Signaling by SCF-KIT / Constitutive Signaling by EGFRvIII / RAF activation / cellular response to gamma radiation / Signaling by high-kinase activity BRAF mutants / Signaling by ERBB2 ECD mutants / MAP2K and MAPK activation / Signaling by ERBB2 KD Mutants / positive regulation of JNK cascade / cellular senescence / positive regulation of fibroblast proliferation Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.25 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.25 Å | ||||||
 Authors Authors | Fetics, S.K. / Kearney, B.M. / Buhrman, G. / Mattos, C. | ||||||
 Citation Citation | Journal: Structure / Year: 2015 Title: Allosteric Effects of the Oncogenic RasQ61L Mutant on Raf-RBD. Authors: Fetics, S.K. / Guterres, H. / Kearney, B.M. / Buhrman, G. / Ma, B. / Nussinov, R. / Mattos, C. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 4g3x.cif.gz | 110.1 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb4g3x.ent.gz | 84.8 KB | Display | PDB format |
| PDBx/mmJSON format | 4g3x.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/g3/4g3xftp://data.pdbj.org/pub/pdb/validation_reports/g3/4g3x | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  4g0nC  1guaS  3k8yS C: citing same article ( S: Starting model for refinement |
|---|---|
| Similar structure data |








-Links
 PDBj
PDBj
















- Assembly
Assembly
| Deposited unit | 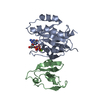
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 18860.221 Da / Num. of mol.: 1 / Mutation: Q61L Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: HRAS, HRAS1 / Plasmid: pET21 / Production host:  |
|---|---|
| #2: Protein | Mass: 8789.290 Da / Num. of mol.: 1 / Fragment: UNP residues 55-131 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: RAF1, RAF / Plasmid: pET302/NT-His / Production host: References: UniProt: P04049, non-specific serine/threonine protein kinase |
| #3: Chemical | ChemComp-GNP / 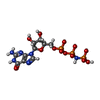  Mass: 522.196 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H17N6O13P3 Mass: 522.196 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H17N6O13P3Comment: GppNHp, GMPPNP, energy-carrying molecule analogue*YM |
| #4: Chemical | ChemComp-MG /   Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg |
| #5: Water | ChemComp-HOH /  Mass: 18.015 Da / Num. of mol.: 17 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 17 / Source method: isolated from a natural source / Formula: H2O |
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 4.06 Å3/Da / Density % sol: 69.71 % |
|---|---|
| Crystal grow | Temperature: 291 K / Method: vapor diffusion, hanging drop / pH: 6.8 Details: Protein solution: 10 - 18 mg/mL, 50 mM HEPES, pH 7.2, 50 mM NaCl, 10mM MgCl2 5% Glycerol, 1mM DTE, 10 M ZnCl2 Reservoir solution:200mM calcium acetate, 100mM sodium cacodylate pH 6.5, 18% ...Details: Protein solution: 10 - 18 mg/mL, 50 mM HEPES, pH 7.2, 50 mM NaCl, 10mM MgCl2 5% Glycerol, 1mM DTE, 10 M ZnCl2 Reservoir solution:200mM calcium acetate, 100mM sodium cacodylate pH 6.5, 18% PEG 8000. Drop: 3uL protein, 3uL reservoir , VAPOR DIFFUSION, HANGING DROP, temperature 291K |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: APS  / Beamline: 22-ID / Wavelength: 1 Å / Beamline: 22-ID / Wavelength: 1 Å |
| Detector | Type: MARMOSAIC 300 mm CCD / Detector: CCD / Date: Jul 1, 2011 / Details: crystal |
| Radiation | Monochromator: crystal / Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1 Å / Relative weight: 1 |
| Reflection | Resolution: 3.25→32.6 Å / Num. all: 6462 / Num. obs: 6462 / Observed criterion σ(F): 3 / Observed criterion σ(I): 3 / Redundancy: 4.6 % / Rmerge(I) obs: 0.18 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: PDB 3K8Y for Chain A and PDB 1GUA for Chain B Resolution: 3.25→32.6 Å / σ(F): 0 / Phase error: 32.62 / Stereochemistry target values: TWIN_LSQ_F
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 3.25→32.6 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|