 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-3j95: Structure of ADP-bound N-ethylmaleimide sensitive factor determin... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 3j95 | ||||||
|---|---|---|---|---|---|---|---|
| Title | Structure of ADP-bound N-ethylmaleimide sensitive factor determined by single particle cryoelectron microscopy | ||||||
 Components Components | Vesicle-fusing ATPase | ||||||
 Keywords Keywords | HYDROLASE / ATPases associated with diverse cellular activities | ||||||
| Function / homology |  Function and homology information Function and homology informationSNARE complex disassembly / ATP-dependent protein disaggregase activity / intra-Golgi vesicle-mediated transport / Golgi to plasma membrane protein transport / Golgi stack / syntaxin-1 binding / vesicle-fusing ATPase / positive regulation of receptor recycling / ionotropic glutamate receptor binding / SNARE binding ...SNARE complex disassembly / ATP-dependent protein disaggregase activity / intra-Golgi vesicle-mediated transport / Golgi to plasma membrane protein transport / Golgi stack / syntaxin-1 binding / vesicle-fusing ATPase / positive regulation of receptor recycling / ionotropic glutamate receptor binding / SNARE binding / PDZ domain binding / intracellular protein transport / positive regulation of protein catabolic process / midbody / protein kinase binding / protein-containing complex binding / ATP hydrolysis activity / ATP binding / metal ion binding / identical protein binding / plasma membrane / cytosol Similarity search - Function | ||||||
| Biological species |   Cricetulus griseus (Chinese hamster) Cricetulus griseus (Chinese hamster) | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 7.6 Å | ||||||
 Authors Authors | Zhao, M. / Wu, S. / Cheng, Y. / Brunger, A.T. | ||||||
 Citation Citation | Journal: Nature / Year: 2015 Title: Mechanistic insights into the recycling machine of the SNARE complex. Authors: Minglei Zhao / Shenping Wu / Qiangjun Zhou / Sandro Vivona / Daniel J Cipriano / Yifan Cheng / Axel T Brunger /  Abstract: Evolutionarily conserved SNARE (soluble N-ethylmaleimide sensitive factor attachment protein receptors) proteins form a complex that drives membrane fusion in eukaryotes. The ATPase NSF (N- ...Evolutionarily conserved SNARE (soluble N-ethylmaleimide sensitive factor attachment protein receptors) proteins form a complex that drives membrane fusion in eukaryotes. The ATPase NSF (N-ethylmaleimide sensitive factor), together with SNAPs (soluble NSF attachment protein), disassembles the SNARE complex into its protein components, making individual SNAREs available for subsequent rounds of fusion. Here we report structures of ATP- and ADP-bound NSF, and the NSF/SNAP/SNARE (20S) supercomplex determined by single-particle electron cryomicroscopy at near-atomic to sub-nanometre resolution without imposing symmetry. Large, potentially force-generating, conformational differences exist between ATP- and ADP-bound NSF. The 20S supercomplex exhibits broken symmetry, transitioning from six-fold symmetry of the NSF ATPase domains to pseudo four-fold symmetry of the SNARE complex. SNAPs interact with the SNARE complex with an opposite structural twist, suggesting an unwinding mechanism. The interfaces between NSF, SNAPs, and SNAREs exhibit characteristic electrostatic patterns, suggesting how one NSF/SNAP species can act on many different SNARE complexes. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 3j95.cif.gz | 483.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb3j95.ent.gz | 361.3 KB | Display | PDB format |
| PDBx/mmJSON format | 3j95.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/j9/3j95ftp://data.pdbj.org/pub/pdb/validation_reports/j9/3j95 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  6205MC 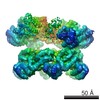 6204C 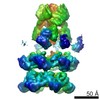 6206C 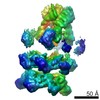 6207C 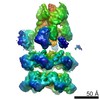 6208C  6209C  6210C  3j94C  3j96C  3j97C  3j98C  3j99C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj

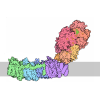

- Assembly
Assembly
| Deposited unit | 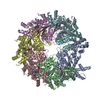
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 82907.430 Da / Num. of mol.: 6 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Cricetulus griseus (Chinese hamster) / Gene: NSF / Production host:  #2: Chemical | ChemComp-ADP /   Mass: 427.201 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: ADP-bound N-ethylmaleimide sensitive factor / Type: COMPLEX / Details: hexamer |
|---|---|
| Molecular weight | Value: 0.5 MDa / Experimental value: NO |
| Buffer solution | Name: 50 mM Tris-Cl, 150 mM NaCl, 1 mM EDTA, 1 mM ATP, 1 mM DTT, 0.05% v/v Nonident P-40 pH: 8 Details: 50 mM Tris-Cl, 150 mM NaCl, 1 mM EDTA, 1 mM ATP, 1 mM DTT, 0.05% v/v Nonident P-40 |
| Specimen | Conc.: 15 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Details: Holey carbon on top of 400 mesh copper grid |
| Vitrification | Instrument: FEI VITROBOT MARK I / Cryogen name: ETHANE / Temp: 90 K / Humidity: 100 % Details: Blot for 3.5 seconds before plunging into liquid ethane (FEI VITROBOT MARK I). Method: Blot for 3.5 seconds before plunging |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Tecnai Polara / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI POLARA 300 / Date: Jan 14, 2014 |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 31000 X / Nominal defocus max: -2800 nm / Nominal defocus min: -1800 nm / Cs: 2.3 mm / Camera length: 0 mm |
| Specimen holder | Specimen holder model: OTHER / Specimen holder type: unspecified |
| Image recording | Electron dose: 44 e/Å2 / Film or detector model: GATAN K2 (4k x 4k) |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Relative weight: 1 |
- Processing
Processing
| EM software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Details: Each particle | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 7.6 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 12830 / Nominal pixel size: 2.4312 Å / Actual pixel size: 2.4312 Å / Details: (Single particle--Applied symmetry: C1) / Symmetry type: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: RECIPROCAL / Target criteria: R-factor Details: REFINEMENT PROTOCOL--flexible DETAILS--D2 domain of NSF was from crystal structure 1NSF. D1 domain of NSF was from related entry EMD-6204. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | PDB-ID: 1NSF Pdb chain-ID: A / Accession code: 1NSF / Source name: PDB / Type: experimental model | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Resolution: 7.6→7.6 Å / SU ML: 1.3 / σ(F): 0.29 / Phase error: 36.78 / Stereochemistry target values: ML
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso max: 50 Å2 / Biso mean: 50 Å2 / Biso min: 50 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 7.601→311.194 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Refine-ID: ELECTRON MICROSCOPY / Total num. of bins used: 30
|

