 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-3aae: Crystal structure of Actin capping protein in complex with CARMIL... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 3aae | ||||||
|---|---|---|---|---|---|---|---|
| Title | Crystal structure of Actin capping protein in complex with CARMIL fragment | ||||||
 Components Components |
| ||||||
 Keywords Keywords | PROTEIN BINDING / ACTIN CAPPING PROTEIN / BARBED END REGULATION / CARMIL FAMILY PROTEIN / CONFORMATIONAL CHANGE / CELL MOTILITY / Actin capping / Actin-binding / Cytoskeleton / Isopeptide bond / Leucine-rich repeat | ||||||
| Function / homology |  Function and homology information Function and homology informationbarbed-end actin filament uncapping / positive regulation of lamellipodium organization / negative regulation of barbed-end actin filament capping / Advanced glycosylation endproduct receptor signaling / : / RHOF GTPase cycle / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / COPI-independent Golgi-to-ER retrograde traffic / macropinosome / Factors involved in megakaryocyte development and platelet production ...barbed-end actin filament uncapping / positive regulation of lamellipodium organization / negative regulation of barbed-end actin filament capping / Advanced glycosylation endproduct receptor signaling / : / RHOF GTPase cycle / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / COPI-independent Golgi-to-ER retrograde traffic / macropinosome / Factors involved in megakaryocyte development and platelet production / COPI-mediated anterograde transport / negative regulation of filopodium assembly / actin filament network formation / F-actin capping protein complex / WASH complex / macropinocytosis / regulation of Arp2/3 complex-mediated actin nucleation / Factors involved in megakaryocyte development and platelet production / urate metabolic process / cell junction assembly / ruffle organization / barbed-end actin filament capping / actin polymerization or depolymerization / regulation of lamellipodium assembly / intermediate filament cytoskeleton / regulation of cell morphogenesis / lamellipodium assembly / filamentous actin / positive regulation of actin filament polymerization / cell leading edge / cortical cytoskeleton / establishment or maintenance of cell polarity / brush border / sperm head-tail coupling apparatus / positive regulation of stress fiber assembly / cytoskeleton organization / positive regulation of substrate adhesion-dependent cell spreading / Schaffer collateral - CA1 synapse / cell morphogenesis / Z disc / actin filament binding / cell migration / lamellipodium / actin cytoskeleton organization / nuclear speck / postsynaptic density / positive regulation of cell migration / protein-containing complex binding / membrane / nucleus / plasma membrane / cytoplasm / cytosol Similarity search - Function | ||||||
| Biological species |   | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.3 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.3 Å | ||||||
 Authors Authors | Takeda, S. / Minakata, S. / Narita, A. / Kitazawa, M. / Yamakuni, T. / Maeda, Y. / Nitanai, Y. | ||||||
 Citation Citation | Journal: To be Published Title: Two distinct mechanisms for the regulation of actin capping protein-competitive or allosteric inhibitions Authors: Takeda, S. / Minakata, S. / Narita, A. / Kitazawa, M. / Yamakuni, T. / Maeda, Y. / Nitanai, Y. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 3aae.cif.gz | 536.2 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb3aae.ent.gz | 444.7 KB | Display | PDB format |
| PDBx/mmJSON format | 3aae.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/aa/3aaeftp://data.pdbj.org/pub/pdb/validation_reports/aa/3aae | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  3aa0S S: Starting model for refinement |
|---|---|
| Similar structure data |
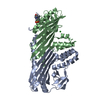
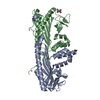






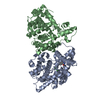
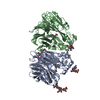

-Links
 PDBj
PDBj






- Assembly
Assembly
| Deposited unit | 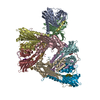
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 | 
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3 | 
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4 | 
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5 | 
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Unit cell |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Noncrystallographic symmetry (NCS) | NCS domain:
NCS domain segments:
NCS ensembles :
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Details | THE ORIGOMETRIC STATE OF THE ACTIN CAPPING PROTEIN IS A HETERO DIMER COMPOSE OF SUBUNIT A (CHAIN A) AND SUBUNIT B (CHAIN B) IN VIVO AND IN VITRO. |
-Components
| #1: Protein | Mass: 33001.789 Da / Num. of mol.: 5 Source method: isolated from a genetically manipulated source Source: (gene. exp.)  #2: Protein | Mass: 31403.449 Da / Num. of mol.: 5 Source method: isolated from a genetically manipulated source Source: (gene. exp.) #3: Protein/peptide | Mass: 4167.751 Da / Num. of mol.: 5 / Fragment: CP-BINDING MOTIF, residues 971-1002 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.87 Å3/Da / Density % sol: 57.12 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, hanging drop / pH: 6.5 Details: 14% PEG 3350, 180MM TRI-AMMONIUM CITRATE, 100MM MES-NAOH, pH 6.5, VAPOR DIFFUSION, HANGING DROP, temperature 293K |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: SPring-8  / Beamline: BL44B2 / Wavelength: 1 Å / Beamline: BL44B2 / Wavelength: 1 Å |
| Detector | Type: ADSC QUANTUM 4r / Detector: CCD / Date: Apr 23, 2008 / Details: mirrors |
| Radiation | Monochromator: SI / Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1 Å / Relative weight: 1 |
| Reflection | Resolution: 3.3→49.7 Å / Num. obs: 58413 / % possible obs: 99.5 % / Redundancy: 7.6 % / Biso Wilson estimate: 38.1 Å2 / Rmerge(I) obs: 0.126 / Net I/σ(I): 16.4 |
| Reflection shell | Resolution: 3.3→3.42 Å / Redundancy: 7.7 % / Rmerge(I) obs: 0.382 / Mean I/σ(I) obs: 5.56 / Num. unique all: 5823 / % possible all: 99.3 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: PDB ENTRY 3AA0 Resolution: 3.3→49.7 Å / Cor.coef. Fo:Fc: 0.869 / Cor.coef. Fo:Fc free: 0.843 / Occupancy max: 1 / Occupancy min: 1 / SU B: 27.301 / SU ML: 0.45 / Cross valid method: THROUGHOUT / σ(F): 0 / ESU R Free: 0.576 / Stereochemistry target values: MAXIMUM LIKELIHOOD / Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.4 Å / Solvent model: MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso max: 99.82 Å2 / Biso mean: 50.947 Å2 / Biso min: 27.63 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine analyze | Luzzati coordinate error obs: 0.67 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 3.3→49.7 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints NCS | Dom-ID: 1 / Refine-ID: X-RAY DIFFRACTION
|