 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 1q4k | ||||||
|---|---|---|---|---|---|---|---|
| Title | The polo-box domain of Plk1 in complex with a phospho-peptide | ||||||
 Components Components |
| ||||||
 Keywords Keywords | TRANSFERASE / Six-stranded anti-parallel beta sheet with one alpha helix | ||||||
| Function / homology |  Function and homology information Function and homology informationpositive regulation of mitotic nuclear envelope disassembly / Mitotic Telophase/Cytokinesis / regulation of protein localization to cell cortex / Mitotic Metaphase/Anaphase Transition / synaptonemal complex disassembly / Activation of NIMA Kinases NEK9, NEK6, NEK7 / polo kinase / Phosphorylation of Emi1 / homologous chromosome segregation / mitotic nuclear membrane disassembly ...positive regulation of mitotic nuclear envelope disassembly / Mitotic Telophase/Cytokinesis / regulation of protein localization to cell cortex / Mitotic Metaphase/Anaphase Transition / synaptonemal complex disassembly / Activation of NIMA Kinases NEK9, NEK6, NEK7 / polo kinase / Phosphorylation of Emi1 / homologous chromosome segregation / mitotic nuclear membrane disassembly / metaphase/anaphase transition of mitotic cell cycle / female meiosis chromosome segregation / anaphase-promoting complex binding / synaptonemal complex / Phosphorylation of the APC/C / astral microtubule organization / Golgi inheritance / mitotic cleavage furrow formation / outer kinetochore / microtubule bundle formation / mitotic chromosome condensation / double-strand break repair via alternative nonhomologous end joining / regulation of mitotic spindle assembly / Polo-like kinase mediated events / Golgi Cisternae Pericentriolar Stack Reorganization / positive regulation of mitotic metaphase/anaphase transition / centrosome cycle / positive regulation of ubiquitin-dependent protein catabolic process / regulation of mitotic metaphase/anaphase transition / sister chromatid cohesion / mitotic spindle pole / mitotic spindle assembly checkpoint signaling / regulation of mitotic cell cycle phase transition / spindle midzone / regulation of anaphase-promoting complex-dependent catabolic process / mitotic G2 DNA damage checkpoint signaling / mitotic cytokinesis / mitotic sister chromatid segregation / negative regulation of double-strand break repair via homologous recombination / Regulation of MITF-M-dependent genes involved in cell cycle and proliferation / Cyclin A/B1/B2 associated events during G2/M transition / protein localization to chromatin / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / Recruitment of mitotic centrosome proteins and complexes / Recruitment of NuMA to mitotic centrosomes / Anchoring of the basal body to the plasma membrane / regulation of mitotic cell cycle / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / AURKA Activation by TPX2 / Resolution of Sister Chromatid Cohesion / mitotic spindle organization / Condensation of Prophase Chromosomes / regulation of cytokinesis / establishment of protein localization / centriole / RHO GTPases Activate Formins / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / positive regulation of protein localization to nucleus / kinetochore / G2/M transition of mitotic cell cycle / spindle / centriolar satellite / spindle pole / The role of GTSE1 in G2/M progression after G2 checkpoint / Separation of Sister Chromatids / Regulation of PLK1 Activity at G2/M Transition / double-strand break repair / mitotic cell cycle / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / microtubule cytoskeleton / midbody / microtubule binding / protein phosphorylation / protein kinase activity / regulation of cell cycle / protein serine kinase activity / protein serine/threonine kinase activity / centrosome / protein kinase binding / negative regulation of apoptotic process / chromatin / magnesium ion binding / negative regulation of transcription by RNA polymerase II / nucleoplasm / ATP binding / identical protein binding / nucleus / cytoplasm / cytosol Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.3 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.3 Å | ||||||
 Authors Authors | Cheng, K. / Lowe, E.D. / Sinclair, J. / Nigg, E.A. / Johnson, L.N. | ||||||
 Citation Citation | Journal: Embo J. / Year: 2003 Title: The crystal structure of the human polo-like kinase-1 polo box domain and its phospho-peptide complex. Authors: Cheng, K.Y. / Lowe, E.D. / Sinclair, J. / Nigg, E.A. / Johnson, L.N. | ||||||
| History |
| ||||||
| Remark 999 | SEQUENCE No sequence database reference found for phospho-peptide (entity 2, chains D,E,F)at the ...SEQUENCE No sequence database reference found for phospho-peptide (entity 2, chains D,E,F)at the time of processing |
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 1q4k.cif.gz | 151 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb1q4k.ent.gz | 119.3 KB | Display | PDB format |
| PDBx/mmJSON format | 1q4k.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/q4/1q4kftp://data.pdbj.org/pub/pdb/validation_reports/q4/1q4k | HTTPS FTP |
|---|
-Related structure data











-Links
 PDBj
PDBj







- Assembly
Assembly
| Deposited unit | 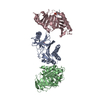
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| 3 | 
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 29862.035 Da / Num. of mol.: 3 / Fragment: Polo box domain of human Plk1 / Mutation: residues 345-603 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PLK1 / Plasmid: pGEX-6P1 / Production host:  References: UniProt: P53350, Transferases; Transferring phosphorus-containing groups; Phosphotransferases with an alcohol group as acceptor #2: Protein/peptide | Mass: 755.774 Da / Num. of mol.: 3 / Fragment: Phospho-peptide / Mutation: sequence Met.Gln.Ser.pThr.Pro.Leu / Source method: obtained synthetically / Details: Chemical synthesis #3: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 191 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 191 / Source method: isolated from a natural source / Formula: H2OHas protein modification | Y | |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.54 Å3/Da / Density % sol: 51.56 % | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, sitting drop / pH: 6.5 Details: PEG 20000, MES, pH 6.5, VAPOR DIFFUSION, SITTING DROP, temperature 293K | ||||||||||||||||||||||||
| Crystal grow | *PLUS Method: vapor diffusion, sitting drop | ||||||||||||||||||||||||
| Components of the solutions | *PLUS
|
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: ESRF  / Beamline: ID29 / Wavelength: 0.9792 Å / Beamline: ID29 / Wavelength: 0.9792 Å |
| Detector | Type: ADSC QUANTUM 4 / Detector: CCD / Date: Jul 6, 2003 / Details: mirrors |
| Radiation | Monochromator: Si monochromator / Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.9792 Å / Relative weight: 1 |
| Reflection | Resolution: 2.3→29.36 Å / Num. all: 38616 / Num. obs: 37228 / % possible obs: 95.7 % / Observed criterion σ(F): 0 / Observed criterion σ(I): 0 / Redundancy: 1.9 % / Biso Wilson estimate: 49.7 Å2 / Rmerge(I) obs: 0.068 / Rsym value: 0.096 / Net I/σ(I): 2.7 |
| Reflection shell | Resolution: 2.3→2.42 Å / Redundancy: 1.9 % / Rmerge(I) obs: 0.364 / Mean I/σ(I) obs: 2.1 / Num. unique all: 5668 / Rsym value: 0.258 / % possible all: 96.6 |
| Reflection | *PLUS Lowest resolution: 24.7 Å / Num. obs: 35628 / Num. measured all: 71256 / Rmerge(I) obs: 0.065 |
| Reflection shell | *PLUS % possible obs: 96.6 % / Rmerge(I) obs: 0.258 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: Unliganded polo box domain Resolution: 2.3→29.36 Å / Cor.coef. Fo:Fc: 0.909 / Cor.coef. Fo:Fc free: 0.861 / SU B: 7.791 / SU ML: 0.196 / Cross valid method: THROUGHOUT / σ(F): 0 / ESU R: 0.373 / ESU R Free: 0.291 / Stereochemistry target values: MAXIMUM LIKELIHOOD / Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.4 Å / Solvent model: BABINET MODEL WITH MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 44.646 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.3→29.36 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Resolution: 2.3→2.359 Å / Total num. of bins used: 20 /
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | *PLUS Lowest resolution: 24.7 Å / Rfactor Rfree: 0.313 / Rfactor Rwork: 0.244 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | *PLUS | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | *PLUS | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints | *PLUS
|
