positive regulation of mitotic nuclear envelope disassembly / Mitotic Telophase/Cytokinesis / regulation of protein localization to cell cortex / Mitotic Metaphase/Anaphase Transition / Activation of NIMA Kinases NEK9, NEK6, NEK7 / polo kinase / Phosphorylation of Emi1 / mitotic nuclear membrane disassembly / metaphase/anaphase transition of mitotic cell cycle / anaphase-promoting complex binding ...positive regulation of mitotic nuclear envelope disassembly / Mitotic Telophase/Cytokinesis / regulation of protein localization to cell cortex / Mitotic Metaphase/Anaphase Transition / Activation of NIMA Kinases NEK9, NEK6, NEK7 / polo kinase / Phosphorylation of Emi1 / mitotic nuclear membrane disassembly / metaphase/anaphase transition of mitotic cell cycle / anaphase-promoting complex binding / synaptonemal complex / Phosphorylation of the APC/C / astral microtubule organization / Golgi inheritance / mitotic cleavage furrow formation / outer kinetochore / microtubule bundle formation / mitotic chromosome condensation / double-strand break repair via alternative nonhomologous end joining / regulation of mitotic spindle assembly / Polo-like kinase mediated events / Golgi Cisternae Pericentriolar Stack Reorganization / positive regulation of mitotic metaphase/anaphase transition / positive regulation of ubiquitin-dependent protein catabolic process / centrosome cycle / regulation of mitotic metaphase/anaphase transition / sister chromatid cohesion / mitotic spindle assembly checkpoint signaling / mitotic spindle pole / regulation of mitotic cell cycle phase transition / spindle midzone / mitotic G2 DNA damage checkpoint signaling / regulation of anaphase-promoting complex-dependent catabolic process / mitotic cytokinesis / mitotic sister chromatid segregation / negative regulation of double-strand break repair via homologous recombination / Regulation of MITF-M-dependent genes involved in cell cycle and proliferation / Cyclin A/B1/B2 associated events during G2/M transition / centriolar satellite / protein localization to chromatin / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / Recruitment of mitotic centrosome proteins and complexes / Recruitment of NuMA to mitotic centrosomes / centriole / Anchoring of the basal body to the plasma membrane / regulation of mitotic cell cycle / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / AURKA Activation by TPX2 / Resolution of Sister Chromatid Cohesion / mitotic spindle organization / Condensation of Prophase Chromosomes / regulation of cytokinesis / establishment of protein localization / RHO GTPases Activate Formins / positive regulation of protein localization to nucleus / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / G2/M transition of mitotic cell cycle / kinetochore / spindle / spindle pole / The role of GTSE1 in G2/M progression after G2 checkpoint / Separation of Sister Chromatids / Regulation of PLK1 Activity at G2/M Transition / double-strand break repair / mitotic cell cycle / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / microtubule cytoskeleton / midbody / microtubule binding / protein phosphorylation / protein kinase activity / regulation of cell cycle / protein serine kinase activity / protein serine/threonine kinase activity / centrosome / protein kinase binding / negative regulation of apoptotic process / chromatin / magnesium ion binding / negative regulation of transcription by RNA polymerase II / nucleoplasm / ATP binding / identical protein binding / nucleus / cytosol / cytoplasm Similarity search - Function
POLO box domain / Polo-like kinase 1, catalytic domain / Second polo-box domain / First polo-box domain / POLO box domain superfamily / POLO box duplicated region / Arylsulfatase, C-terminal domain / POLO box domain / POLO box domain profile. / Serine/threonine-protein kinase, active site ...POLO box domain / Polo-like kinase 1, catalytic domain / Second polo-box domain / First polo-box domain / POLO box domain superfamily / POLO box duplicated region / Arylsulfatase, C-terminal domain / POLO box domain / POLO box domain profile. / Serine/threonine-protein kinase, active site / Serine/Threonine protein kinases active-site signature. / Protein kinase domain / Serine/Threonine protein kinases, catalytic domain / Protein kinase, ATP binding site / Protein kinases ATP-binding region signature. / Protein kinase domain profile. / Protein kinase domain / Protein kinase-like domain superfamily / 2-Layer Sandwich / Alpha Beta Similarity search - Domain/homology
Resolution: 2.75→51.07 Å / Cor.coef. Fo:Fc: 0.897 / Cor.coef. Fo:Fc free: 0.797 / SU B: 18.386 / SU ML: 0.375 / Cross valid method: THROUGHOUT / ESU R Free: 0.469 / Stereochemistry target values: MAXIMUM LIKELIHOOD / Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
Rfactor
Num. reflection
% reflection
Selection details
Rfree
0.2802
531
4.8 %
RANDOM
Rwork
0.20588
-
-
-
obs
0.20956
10576
94.18 %
-
Solvent computation
Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.4 Å / Solvent model: MASK
Displacement parameters
Biso mean: 15.541 Å2
Baniso -1
Baniso -2
Baniso -3
1-
-0.11 Å2
0 Å2
0.04 Å2
2-
-
0.04 Å2
0 Å2
3-
-
-
0.07 Å2
Refinement step
Cycle: LAST / Resolution: 2.75→51.07 Å
Protein
Nucleic acid
Ligand
Solvent
Total
Num. atoms
3519
0
18
103
3640
Refine LS restraints
Refine-ID
Type
Dev ideal
Dev ideal target
Number
X-RAY DIFFRACTION
r_bond_refined_d
0.008
0.022
3609
X-RAY DIFFRACTION
r_bond_other_d
X-RAY DIFFRACTION
r_angle_refined_deg
1.126
1.974
4881
X-RAY DIFFRACTION
r_angle_other_deg
X-RAY DIFFRACTION
r_dihedral_angle_1_deg
5.742
5
436
X-RAY DIFFRACTION
r_dihedral_angle_2_deg
29.402
22.988
164
X-RAY DIFFRACTION
r_dihedral_angle_3_deg
16.758
15
628
X-RAY DIFFRACTION
r_dihedral_angle_4_deg
20.312
15
29
X-RAY DIFFRACTION
r_chiral_restr
0.076
0.2
543
X-RAY DIFFRACTION
r_gen_planes_refined
0.004
0.021
2699
X-RAY DIFFRACTION
r_gen_planes_other
X-RAY DIFFRACTION
r_nbd_refined
X-RAY DIFFRACTION
r_nbd_other
X-RAY DIFFRACTION
r_nbtor_refined
X-RAY DIFFRACTION
r_nbtor_other
X-RAY DIFFRACTION
r_xyhbond_nbd_refined
X-RAY DIFFRACTION
r_xyhbond_nbd_other
X-RAY DIFFRACTION
r_metal_ion_refined
X-RAY DIFFRACTION
r_metal_ion_other
X-RAY DIFFRACTION
r_symmetry_vdw_refined
X-RAY DIFFRACTION
r_symmetry_vdw_other
X-RAY DIFFRACTION
r_symmetry_hbond_refined
X-RAY DIFFRACTION
r_symmetry_hbond_other
X-RAY DIFFRACTION
r_symmetry_metal_ion_refined
X-RAY DIFFRACTION
r_symmetry_metal_ion_other
X-RAY DIFFRACTION
r_mcbond_it
0.374
1.5
2187
X-RAY DIFFRACTION
r_mcbond_other
X-RAY DIFFRACTION
r_mcangle_it
0.711
2
3515
X-RAY DIFFRACTION
r_scbond_it
0.896
3
1422
X-RAY DIFFRACTION
r_scangle_it
1.511
4.5
1366
X-RAY DIFFRACTION
r_rigid_bond_restr
X-RAY DIFFRACTION
r_sphericity_free
X-RAY DIFFRACTION
r_sphericity_bonded
LS refinement shell
Resolution: 2.75→2.821 Å / Total num. of bins used: 20
Rfactor
Num. reflection
% reflection
Rfree
0.435
33
-
Rwork
0.286
754
-
obs
-
-
90.25 %
+
About Yorodumi
-
News
-
Feb 9, 2022. New format data for meta-information of EMDB entries
New format data for meta-information of EMDB entries
Version 3 of the EMDB header file is now the official format.
The previous official version 1.9 will be removed from the archive.
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller

 Yorodumi
Yorodumi Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information Homo sapiens (human)
Homo sapiens (human) X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads










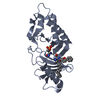
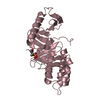
 PDBj
PDBj







 Assembly
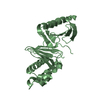
Assembly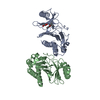


 Mass: 92.094 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C3H8O3
Mass: 92.094 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C3H8O3
 Mass: 164.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H12O2
Mass: 164.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H12O2 Mass: 18.015 Da / Num. of mol.: 103 / Source method: isolated from a natural source / Formula: H2O
Mass: 18.015 Da / Num. of mol.: 103 / Source method: isolated from a natural source / Formula: H2O Sample preparation
Sample preparation Processing
Processing