positive regulation of mitotic nuclear envelope disassembly / Mitotic Telophase/Cytokinesis / regulation of protein localization to cell cortex / Mitotic Metaphase/Anaphase Transition / Activation of NIMA Kinases NEK9, NEK6, NEK7 / polo kinase / Phosphorylation of Emi1 / mitotic nuclear membrane disassembly / metaphase/anaphase transition of mitotic cell cycle / anaphase-promoting complex binding ...positive regulation of mitotic nuclear envelope disassembly / Mitotic Telophase/Cytokinesis / regulation of protein localization to cell cortex / Mitotic Metaphase/Anaphase Transition / Activation of NIMA Kinases NEK9, NEK6, NEK7 / polo kinase / Phosphorylation of Emi1 / mitotic nuclear membrane disassembly / metaphase/anaphase transition of mitotic cell cycle / anaphase-promoting complex binding / synaptonemal complex / Phosphorylation of the APC/C / astral microtubule organization / Golgi inheritance / mitotic cleavage furrow formation / outer kinetochore / microtubule bundle formation / mitotic chromosome condensation / double-strand break repair via alternative nonhomologous end joining / regulation of mitotic spindle assembly / Polo-like kinase mediated events / Golgi Cisternae Pericentriolar Stack Reorganization / positive regulation of mitotic metaphase/anaphase transition / positive regulation of ubiquitin-dependent protein catabolic process / centrosome cycle / regulation of mitotic metaphase/anaphase transition / sister chromatid cohesion / mitotic spindle assembly checkpoint signaling / mitotic spindle pole / regulation of mitotic cell cycle phase transition / spindle midzone / mitotic G2 DNA damage checkpoint signaling / regulation of anaphase-promoting complex-dependent catabolic process / mitotic cytokinesis / mitotic sister chromatid segregation / negative regulation of double-strand break repair via homologous recombination / Regulation of MITF-M-dependent genes involved in cell cycle and proliferation / Cyclin A/B1/B2 associated events during G2/M transition / centriolar satellite / protein localization to chromatin / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / Recruitment of mitotic centrosome proteins and complexes / Recruitment of NuMA to mitotic centrosomes / centriole / Anchoring of the basal body to the plasma membrane / regulation of mitotic cell cycle / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / AURKA Activation by TPX2 / Resolution of Sister Chromatid Cohesion / mitotic spindle organization / Condensation of Prophase Chromosomes / regulation of cytokinesis / establishment of protein localization / RHO GTPases Activate Formins / positive regulation of protein localization to nucleus / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / G2/M transition of mitotic cell cycle / kinetochore / spindle / spindle pole / The role of GTSE1 in G2/M progression after G2 checkpoint / Separation of Sister Chromatids / Regulation of PLK1 Activity at G2/M Transition / double-strand break repair / mitotic cell cycle / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / microtubule cytoskeleton / midbody / microtubule binding / protein phosphorylation / protein kinase activity / regulation of cell cycle / protein serine kinase activity / protein serine/threonine kinase activity / centrosome / protein kinase binding / negative regulation of apoptotic process / chromatin / magnesium ion binding / negative regulation of transcription by RNA polymerase II / nucleoplasm / ATP binding / identical protein binding / nucleus / cytosol / cytoplasm Similarity search - Function
POLO box domain / Polo-like kinase 1, catalytic domain / Second polo-box domain / First polo-box domain / POLO box domain superfamily / POLO box duplicated region / Arylsulfatase, C-terminal domain / POLO box domain / POLO box domain profile. / Serine/threonine-protein kinase, active site ...POLO box domain / Polo-like kinase 1, catalytic domain / Second polo-box domain / First polo-box domain / POLO box domain superfamily / POLO box duplicated region / Arylsulfatase, C-terminal domain / POLO box domain / POLO box domain profile. / Serine/threonine-protein kinase, active site / Serine/Threonine protein kinases active-site signature. / Protein kinase domain / Serine/Threonine protein kinases, catalytic domain / Protein kinase, ATP binding site / Protein kinases ATP-binding region signature. / Protein kinase domain profile. / Protein kinase domain / Protein kinase-like domain superfamily / 2-Layer Sandwich / Alpha Beta Similarity search - Domain/homology
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information HOMO SAPIENS (human)
HOMO SAPIENS (human) X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads
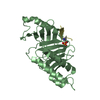






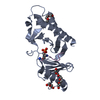

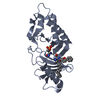

 PDBj
PDBj







 Assembly
Assembly



 Mass: 18.015 Da / Num. of mol.: 275 / Source method: isolated from a natural source / Formula: H2O
Mass: 18.015 Da / Num. of mol.: 275 / Source method: isolated from a natural source / Formula: H2O Sample preparation
Sample preparation / Beamline: PX14.1 / Wavelength: 1.244
/ Beamline: PX14.1 / Wavelength: 1.244  Processing
Processing