 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: EMDB / ID: EMD-4903 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
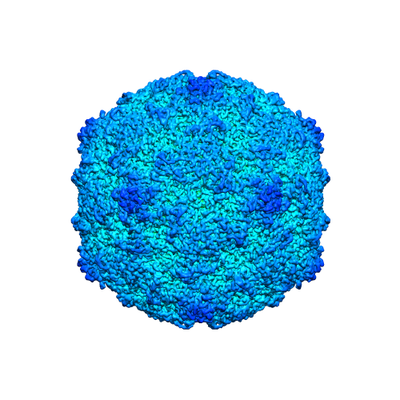
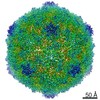
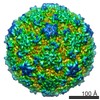
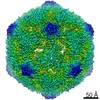
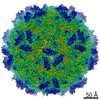
| Title | Echovirus 1 intact particle | |||||||||||||||
 Map data Map data | E1 control | |||||||||||||||
 Sample Sample |
| |||||||||||||||
 Keywords Keywords | VIRAL COAT PROTEIN / CAPSID / PICORNAVIRUS / ECHOVIRUS / ICOSAHEDRAL VIRUS / VIRUS | |||||||||||||||
| Function / homology |  Function and homology information Function and homology informationcaveolin-mediated endocytosis of virus by host cell / symbiont-mediated suppression of host cytoplasmic pattern recognition receptor signaling pathway via inhibition of RIG-I activity / picornain 2A / symbiont-mediated suppression of host mRNA export from nucleus / symbiont genome entry into host cell via pore formation in plasma membrane / picornain 3C / T=pseudo3 icosahedral viral capsid / host cell cytoplasmic vesicle membrane / viral capsid / ribonucleoside triphosphate phosphatase activity ...caveolin-mediated endocytosis of virus by host cell / symbiont-mediated suppression of host cytoplasmic pattern recognition receptor signaling pathway via inhibition of RIG-I activity / picornain 2A / symbiont-mediated suppression of host mRNA export from nucleus / symbiont genome entry into host cell via pore formation in plasma membrane / picornain 3C / T=pseudo3 icosahedral viral capsid / host cell cytoplasmic vesicle membrane / viral capsid / ribonucleoside triphosphate phosphatase activity / nucleoside-triphosphate phosphatase / channel activity / monoatomic ion transmembrane transport / host cell cytoplasm / DNA replication / RNA helicase activity / symbiont-mediated suppression of host gene expression / symbiont-mediated activation of host autophagy / RNA-directed RNA polymerase / cysteine-type endopeptidase activity / viral RNA genome replication / RNA-directed RNA polymerase activity / virion attachment to host cell / host cell nucleus / DNA-templated transcription / structural molecule activity / proteolysis / RNA binding / zinc ion binding / ATP binding / cytoplasm Similarity search - Function | |||||||||||||||
| Biological species |  Echovirus E1 Echovirus E1 | |||||||||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 3.5 Å | |||||||||||||||
 Authors Authors | Domanska A / Ruokolainen VP | |||||||||||||||
| Funding support |  Finland, 4 items Finland, 4 items
| |||||||||||||||
 Citation Citation | Journal: Acta Crystallogr D Biol Crystallogr / Year: 1996 Title: A pseudo-cell based approach to efficient crystallographic refinement of viruses. Authors: D H Jacobson / J M Hogle / D J Filman /  Abstract: Strategies have been developed for the inexpensive refinement of atomic models of viruses and of other highly symmetric structures. These methods, which have been used in the refinement of several ...Strategies have been developed for the inexpensive refinement of atomic models of viruses and of other highly symmetric structures. These methods, which have been used in the refinement of several strains of poliovirus, focus on an arbitrary-sized parallelepiped (termed the 'protomer' box) containing a single complete averaged copy of the structural motif which forms the protein capsid, together with the fragments of other symmetry-related copies of the motif which are located in its immediate neighborhood. The Fourier transform of the protomer box provides reference structure factors for stereochemically restrained crystallographic refinement of the atomic model parameters. The phases of the reference structure factors are based on the averaged map, and are not permitted to change during the refinement. It is demonstrated that models refined using the protomer box methods do not differ significantly from models refined by more expensive full-cell calculations. | |||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | EM map: SurfViewMolmilJmol/JSmol |
| Supplemental images |
- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_4903.map.gz | 221.2 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-4903-v30.xmlemd-4903.xml | 23.1 KB 23.1 KB | Display Display | EMDB header |
| Images | 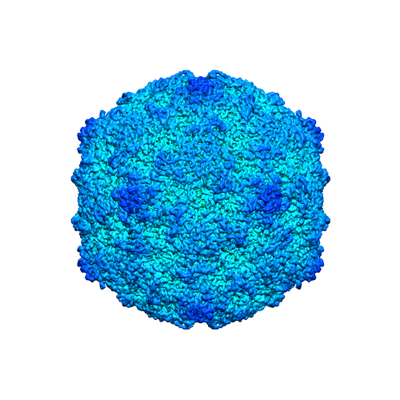 emd_4903.png emd_4903.png | 134.8 KB | ||
| Filedesc metadata | emd-4903.cif.gz | 6.8 KB | ||
| Others | emd_4903_half_map_1.map.gzemd_4903_half_map_2.map.gz | 192.1 MB 192.1 MB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-4903ftp://ftp.pdbj.org/pub/emdb/structures/EMD-4903 http://ftp.pdbj.org/pub/emdb/structures/EMD-4903ftp://ftp.pdbj.org/pub/emdb/structures/EMD-4903 | HTTPS FTP |
-Related structure data
| Related structure data |  6rjfMC  0565C  6o06C M: atomic model generated by this map C: citing same article ( |
|---|---|
| Similar structure data | |
| EM raw data | EMPIAR-10284 (Title: Extracellular albumin and endosomal ions prime enterovirus particles for uncoating that can be prevented by fatty acid saturation Data size: 2.4 TB Data #1: Unaligned multi-frame micrographs of control Echovirus 1 [micrographs - multiframe] Data #2: Unaligned multi-frame micrographs of control Echovirus 1 [micrographs - multiframe] Data #3: Unaligned multi-frame micrographs of treated Echovirus 1 (expanded particle) [micrographs - multiframe]) |



-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| Related items in Molecule of the Month |


-Map
| File | Download / File: emd_4903.map.gz / Format: CCP4 / Size: 244.1 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annotation | E1 control | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 1.24 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Density |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
CCP4 map header:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-Supplemental data
-Half map: E1 control half map 2
| File | emd_4903_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annotation | E1 control half map 2 | ||||||||||||
| Projections & Slices |
| ||||||||||||
| Density Histograms |






-Half map: E1 control half map
| File | emd_4903_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annotation | E1 control half map | ||||||||||||
| Projections & Slices |
| ||||||||||||
| Density Histograms |








- Sample components
Sample components
-Entire : Echovirus E1
| Entire | Name: Echovirus E1 |
|---|---|
| Components |
|
-Supramolecule #1: Echovirus E1
| Supramolecule | Name: Echovirus E1 / type: virus / ID: 1 / Parent: 0 / Macromolecule list: #1-#4 / NCBI-ID: 46633 / Sci species name: Echovirus E1 / Virus type: VIRION / Virus isolate: OTHER / Virus enveloped: No / Virus empty: No |
|---|---|
| Host (natural) | Organism:  Homo sapiens (human) Homo sapiens (human) |
| Virus shell | Shell ID: 1 / Name: icosahedral / Diameter: 300.0 Å / T number (triangulation number): 1 |
-Macromolecule #1: VP1
| Macromolecule | Name: VP1 / type: protein_or_peptide / ID: 1 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Echovirus E1 / Strain: Human/Egypt/Farouk/1951 |
| Molecular weight | Theoretical: 31.604373 KDa |
| Sequence | String: GDVQNAVEGA MVRVADTVQT SATNSERVPN LTAVETGHTS QAVPGDTMQT RHVINNHVRS ESTIENFLAR SACVFYLEYK TGTKEDSNS FNNWVITTRR VAQLRRKLEM FTYLRFDMEI TVVITSSQDQ STSQNQNAPV LTHQIMYVPP GGPIPVSVDD Y SWQTSTNP ...String: GDVQNAVEGA MVRVADTVQT SATNSERVPN LTAVETGHTS QAVPGDTMQT RHVINNHVRS ESTIENFLAR SACVFYLEYK TGTKEDSNS FNNWVITTRR VAQLRRKLEM FTYLRFDMEI TVVITSSQDQ STSQNQNAPV LTHQIMYVPP GGPIPVSVDD Y SWQTSTNP SIFWTEGNAP ARMSIPFISI GNAYSNFYDG WSHFSQAGVY GFTTLNNMGQ LFFRHVNKPN PAAITSVARI YF KPKHVRA WVPRPPRLCP YINSTNVNFE PKPVTEVRTN IITT UniProtKB: Genome polyprotein |
-Macromolecule #2: VP2
| Macromolecule | Name: VP2 / type: protein_or_peptide / ID: 2 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Echovirus E1 |
| Molecular weight | Theoretical: 28.87226 KDa |
| Sequence | String: SPTVEECGYS DRVRSITLGN STITTQECAN VVVGYGEWPE YLSDNEATAE DQPTQPDVAT CRFYTLDSVQ WENGSPGWWW KFPDALRDM GLFGQNMYYH YLGRAGYTIH VQCNASKFHQ GCILVVCVPE AEMGSAQTSG VVNYEHISKG EIASRFTTTT T AEDHGVQA ...String: SPTVEECGYS DRVRSITLGN STITTQECAN VVVGYGEWPE YLSDNEATAE DQPTQPDVAT CRFYTLDSVQ WENGSPGWWW KFPDALRDM GLFGQNMYYH YLGRAGYTIH VQCNASKFHQ GCILVVCVPE AEMGSAQTSG VVNYEHISKG EIASRFTTTT T AEDHGVQA AVWNAGMGVG VGNLTIFPHQ WINLRTNNSA TIVMPYVNSV PMDNMYRHHN FTLMIIPFVP LDFSAGASTY VP ITVTVAP MCAEYNGLRL AGHQ UniProtKB: Genome polyprotein |
-Macromolecule #3: VP3
| Macromolecule | Name: VP3 / type: protein_or_peptide / ID: 3 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Echovirus E1 |
| Molecular weight | Theoretical: 26.471074 KDa |
| Sequence | String: GLPTMNTPGS NQFLTSDDFQ SPSAMPQFDV TPEMHIPGEV RNLMEIAEVD SVMPINNDSA AKVSSMEAYR VELSTNTNAG TQVFGFQLN PGAESVMNRT LMGEILNYYA HWSGSIKITF VFCGSAMTTG KFLLSYAPPG AGAPKTRKDA MLGTHVVWDV G LQSSCVLC ...String: GLPTMNTPGS NQFLTSDDFQ SPSAMPQFDV TPEMHIPGEV RNLMEIAEVD SVMPINNDSA AKVSSMEAYR VELSTNTNAG TQVFGFQLN PGAESVMNRT LMGEILNYYA HWSGSIKITF VFCGSAMTTG KFLLSYAPPG AGAPKTRKDA MLGTHVVWDV G LQSSCVLC IPWISQTHYR FVEKDPYTNA GFVTCWYQTS VVSPASNQPK CYMMCMVSAC NDFSVRMLRD TKFIEQTSFY Q UniProtKB: Genome polyprotein |
-Macromolecule #4: VP4
| Macromolecule | Name: VP4 / type: protein_or_peptide / ID: 4 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Echovirus E1 |
| Molecular weight | Theoretical: 7.398131 KDa |
| Sequence | String: GAQVSTQKTG AHETSLSATG NSIIHYTNIN YYKDAASNSA NRQDFTQDPG KFTEPMKDVM IKTLPALN |
-Macromolecule #5: PALMITIC ACID
| Macromolecule | Name: PALMITIC ACID / type: ligand / ID: 5 / Number of copies: 1 / Formula: PLM |
|---|---|
| Molecular weight | Theoretical: 256.424 Da |
| Chemical component information |  ChemComp-PLM: |
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Buffer | pH: 7.2 / Details: 2 mM magnesium chloride in PBS |
|---|---|
| Grid | Model: Quantifoil R2/2 / Support film - Material: CARBON / Support film - topology: HOLEY / Pretreatment - Type: GLOW DISCHARGE |
| Vitrification | Cryogen name: ETHANE / Instrument: HOMEMADE PLUNGER |
- Electron microscopy
Electron microscopy
| Microscope | FEI TALOS ARCTICA |
|---|---|
| Image recording | Film or detector model: FEI FALCON III (4k x 4k) / Number real images: 979 / Average electron dose: 30.0 e/Å2 |
| Electron beam | Acceleration voltage: 200 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD |
| Sample stage | Cooling holder cryogen: NITROGEN |
| Experimental equipment |  Model: Talos Arctica / Image courtesy: FEI Company |
+Image processing
-Atomic model buiding 1
| Initial model | PDB ID: Chain - Source name: PDB / Chain - Initial model type: experimental model |
|---|---|
| Refinement | Protocol: RIGID BODY FIT |
| Output model | PDB-6rjf: |

-Atomic model buiding 2
| Initial model | PDB ID: Chain - Source name: PDB / Chain - Initial model type: experimental model |
|---|---|
| Refinement | Space: REAL / Protocol: FLEXIBLE FIT |
| Output model | PDB-6rjf: |
