Ribosomal protein L35, mitochondrial / : / Large ribosomal subunit protein bL9m N-terminal domain / : / : / MRPL44, double-stranded RNA binding domain / : / MRPL44 dsRNA-binding domain / Large ribosomal subunit protein mL44, endonuclease domain / : ...Ribosomal protein L35, mitochondrial / : / Large ribosomal subunit protein bL9m N-terminal domain / : / : / MRPL44, double-stranded RNA binding domain / : / MRPL44 dsRNA-binding domain / Large ribosomal subunit protein mL44, endonuclease domain / : / Large ribosomal subunit protein bL9m C-terminal domain / Tim44-like domain / Tim44-like domain / Tim44 / : / : / Ribosomal protein L47, mitochondrial / MRP-L47 superfamily, mitochondrial / Mitochondrial 39-S ribosomal protein L47 (MRP-L47) / Phosphatidylethanolamine-binding protein, eukaryotic / Phosphatidylethanolamine-binding protein / Phosphatidylethanolamine-binding protein / PEBP-like superfamily / Threonyl/alanyl tRNA synthetase, class II-like, putative editing domain superfamily / TGS-like / TGS domain profile. / TGS / Peptide chain release factor class I / RF-1 domain / Ribonuclease III, endonuclease domain superfamily / Double stranded RNA-binding domain (dsRBD) profile. / Double-stranded RNA-binding domain / Beta-grasp domain superfamily / NTF2-like domain superfamily / Ribosomal protein L9, N-terminal domain superfamily / Ribosomal protein L9 / Ribosomal protein L9, N-terminal / Ribosomal protein L9, N-terminal domain / Ribosomal protein L9/RNase H1, N-terminal / Ribosomal protein L18 / Ribosomal L18 of archaea, bacteria, mitoch. and chloroplast / Ribosomal protein L16 / Ribosomal protein L30, bacterial-type / : / Ribosomal protein L27 / Ribosomal L27 protein / Ribosomal L32p protein family / Ribosomal protein L19 / Ribosomal protein L19 / Ribosomal protein L19 superfamily / Ribosomal protein L21 / Ribosomal protein L32p / Ribosomal protein L17 / Ribosomal protein L17 superfamily / Ribosomal protein L17 / Ribosomal proteins 50S L24/mitochondrial 39S L24 / Ribosomal protein L21-like / L21-like superfamily / Ribosomal prokaryotic L21 protein / Ribosomal protein L34 / Ribosomal protein L34 / Ribosomal protein L24 / Ribosomal protein L15, bacterial-type / 50S ribosomal protein uL4 / : / Ribosomal protein L10e/L16 / Ribosomal protein L10e/L16 superfamily / Ribosomal protein L16p/L10e / Ribosomal Proteins L2, RNA binding domain / Ribosomal Proteins L2, RNA binding N-terminal domain / Ribosomal Proteins L2, RNA binding domain / Ribosomal protein L24 signature. / Ribosomal protein L2 / Ribosomal protein L24/L26, conserved site / KOW (Kyprides, Ouzounis, Woese) motif. / Ribosomal protein L23 / Ribosomal protein L30, ferredoxin-like fold domain / Ribosomal protein L30p/L7e / Ribosomal protein L25/L23 / Ribosomal protein L30, ferredoxin-like fold domain superfamily / Ribosomal protein L15 / Ribosomal proteins 50S-L15, 50S-L18e, 60S-L27A / Ribosomal protein S11 superfamily / Ribosomal protein L14p/L23e / Ribosomal protein L14P / Ribosomal protein L14 superfamily / Ribosomal protein L14p/L23e / Ribosomal protein L22/L17 / Ribosomal protein L22p/L17e / Ribosomal protein L22/L17 superfamily / Ribosomal protein L26/L24, KOW domain / Ribosomal protein L18e/L15P / Ribosomal L18e/L15P superfamily / Ribosomal protein L4/L1e / Ribosomal protein L4 domain superfamily / Ribosomal protein L4/L1 family / Ribosomal protein L23/L15e core domain superfamily / Translation protein SH3-like domain superfamily / Zinc-binding ribosomal protein / KOW motif 類似検索 - ドメイン・相同性
Large ribosomal subunit protein uL18m / Large ribosomal subunit protein uL18m / Large ribosomal subunit protein uL24m / Large ribosomal subunit protein uL16m / : / Large ribosomal subunit protein bL17m / Large ribosomal subunit protein uL14m / Mitochondrial ribosomal protein L2 / 39S ribosomal protein L15, mitochondrial / Large ribosomal subunit protein bL27m ...Large ribosomal subunit protein uL18m / Large ribosomal subunit protein uL18m / Large ribosomal subunit protein uL24m / Large ribosomal subunit protein uL16m / : / Large ribosomal subunit protein bL17m / Large ribosomal subunit protein uL14m / Mitochondrial ribosomal protein L2 / 39S ribosomal protein L15, mitochondrial / Large ribosomal subunit protein bL27m / Large ribosomal subunit protein mL38 / Large ribosomal subunit protein uL18m / Large ribosomal subunit protein bL21m / : / Mitochondrial ribosomal protein L4 / Large ribosomal subunit protein uL18m / Large ribosomal subunit protein uL29m / Large ribosomal subunit protein mL39 / Large ribosomal subunit protein mL44 / Large ribosomal subunit protein uL30m / Large ribosomal subunit protein bL35m / : / Large ribosomal subunit protein mL45 / Large ribosomal subunit protein bL19m / Large ribosomal subunit protein bL9m / Large ribosomal subunit protein bL32m / Large ribosomal subunit protein uL22m / Large ribosomal subunit protein uL23m / Large ribosomal subunit protein mL62 / Large ribosomal subunit protein bL34m 類似検索 - 構成要素
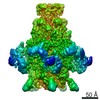
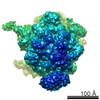
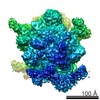
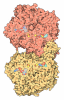
ジャーナル: Nature / 年: 2014 タイトル: Architecture of the large subunit of the mammalian mitochondrial ribosome. 著者: Basil J Greber / Daniel Boehringer / Alexander Leitner / Philipp Bieri / Felix Voigts-Hoffmann / Jan P Erzberger / Marc Leibundgut / Ruedi Aebersold / Nenad Ban / 要旨: Mitochondrial ribosomes synthesize a number of highly hydrophobic proteins encoded on the genome of mitochondria, the organelles in eukaryotic cells that are responsible for energy conversion by ...Mitochondrial ribosomes synthesize a number of highly hydrophobic proteins encoded on the genome of mitochondria, the organelles in eukaryotic cells that are responsible for energy conversion by oxidative phosphorylation. The ribosomes in mammalian mitochondria have undergone massive structural changes throughout their evolution, including ribosomal RNA shortening and acquisition of mitochondria-specific ribosomal proteins. Here we present the three-dimensional structure of the 39S large subunit of the porcine mitochondrial ribosome determined by cryo-electron microscopy at 4.9 Å resolution. The structure, combined with data from chemical crosslinking and mass spectrometry experiments, reveals the unique features of the 39S subunit at near-atomic resolution and provides detailed insight into the architecture of the polypeptide exit site. This region of the mitochondrial ribosome has been considerably remodelled compared to its bacterial counterpart, providing a specialized platform for the synthesis and membrane insertion of the highly hydrophobic protein components of the respiratory chain.
pH: 7.9 / 詳細: 20 mM HEPES-KOH, 100 mM KCl, 1 mM DTT, 20 mM MgCl2
グリッド
詳細: 200 mesh Quantifoil R2/1 and R 2/2 holey carbon grids
凍結
凍結剤: ETHANE / 装置: HOMEMADE PLUNGER
+
電子顕微鏡法 #1
Microscopy ID
1
顕微鏡
FEI TITAN KRIOS
日付
2013年6月21日
撮影
カテゴリ: CCD フィルム・検出器のモデル: FEI FALCON I (4k x 4k) デジタル化 - サンプリング間隔: 14 µm / 平均電子線量: 20 e/Å2 詳細: For data processing Falcon 2 data were resampled to 1.41 Angstrom per pixel
カテゴリ: CCD フィルム・検出器のモデル: FEI FALCON I (4k x 4k) デジタル化 - サンプリング間隔: 14 µm / 平均電子線量: 20 e/Å2 詳細: For data processing Falcon 2 data were resampled to 1.41 Angstrom per pixel
カテゴリ: CCD フィルム・検出器のモデル: FEI FALCON I (4k x 4k) デジタル化 - サンプリング間隔: 14 µm / 平均電子線量: 20 e/Å2 詳細: For data processing Falcon 2 data were resampled to 1.41 Angstrom per pixel
カテゴリ: CCD フィルム・検出器のモデル: FEI FALCON I (4k x 4k) デジタル化 - サンプリング間隔: 14 µm / 平均電子線量: 20 e/Å2 詳細: For data processing Falcon 2 data were resampled to 1.41 Angstrom per pixel
カテゴリ: CCD フィルム・検出器のモデル: FEI FALCON I (4k x 4k) デジタル化 - サンプリング間隔: 14 µm / 平均電子線量: 20 e/Å2 詳細: For data processing Falcon 2 data were resampled to 1.41 Angstrom per pixel
カテゴリ: CCD フィルム・検出器のモデル: FEI FALCON I (4k x 4k) デジタル化 - サンプリング間隔: 14 µm / 平均電子線量: 20 e/Å2 詳細: For data processing Falcon 2 data were resampled to 1.41 Angstrom per pixel
カテゴリ: CCD フィルム・検出器のモデル: FEI FALCON I (4k x 4k) デジタル化 - サンプリング間隔: 14 µm / 平均電子線量: 20 e/Å2 詳細: For data processing Falcon 2 data were resampled to 1.41 Angstrom per pixel
カテゴリ: CCD フィルム・検出器のモデル: FEI FALCON I (4k x 4k) デジタル化 - サンプリング間隔: 14 µm / 平均電子線量: 20 e/Å2 詳細: For data processing Falcon 2 data were resampled to 1.41 Angstrom per pixel
Particles were selected and extracted in batchboxer (EMAN 1.9).
CTF補正
詳細: per detector frame
最終 再構成
想定した対称性 - 点群: C1 (非対称) / アルゴリズム: OTHER / 解像度のタイプ: BY AUTHOR / 解像度: 4.9 Å / 解像度の算出法: OTHER ソフトウェア - 名称: Spider, IMAGIC-5, CTFFIND3, RELION 詳細: An initial reconstruction was obtained using the angular reconstitution approach in Imagic-5. For visualization purposes the final map was amplitude and MTF-corrected in RELION. 使用した粒子像数: 232181
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 引用
引用
 構造の表示
構造の表示 UCSF Chimera
UCSF Chimera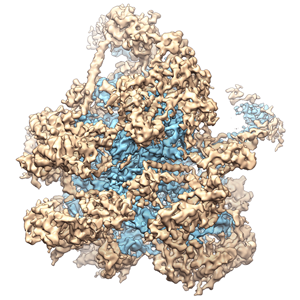
 ダウンロードとリンク
ダウンロードとリンク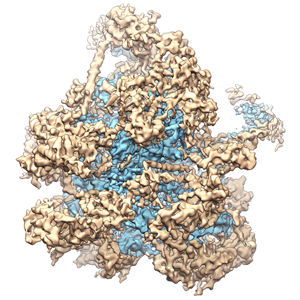 EMD-2490-image_EMDB_ray_500px.png
EMD-2490-image_EMDB_ray_500px.png http://ftp.pdbj.org/pub/emdb/structures/EMD-2490
http://ftp.pdbj.org/pub/emdb/structures/EMD-2490









 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)




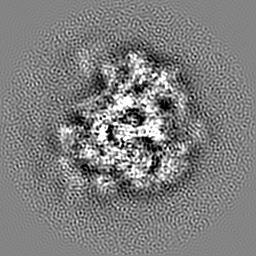















 試料の構成要素
試料の構成要素 解析
解析 電子顕微鏡法 #1
電子顕微鏡法 #1

