piccolo histone acetyltransferase complex / promoter-enhancer loop anchoring activity / telomerase RNA localization to Cajal body / regulation of DNA strand elongation / positive regulation of telomere maintenance in response to DNA damage / sperm DNA condensation / positive regulation of norepinephrine uptake / histone chaperone activity / establishment of protein localization to chromatin / R2TP complex ...piccolo histone acetyltransferase complex / promoter-enhancer loop anchoring activity / telomerase RNA localization to Cajal body / regulation of DNA strand elongation / positive regulation of telomere maintenance in response to DNA damage / sperm DNA condensation / positive regulation of norepinephrine uptake / histone chaperone activity / establishment of protein localization to chromatin / R2TP complex / dynein axonemal particle / bBAF complex / cellular response to cytochalasin B / neural retina development / npBAF complex / nBAF complex / brahma complex / regulation of transepithelial transport / Swr1 complex / Formation of annular gap junctions / morphogenesis of a polarized epithelium / Formation of the dystrophin-glycoprotein complex (DGC) / structural constituent of postsynaptic actin cytoskeleton / GBAF complex / RPAP3/R2TP/prefoldin-like complex / Gap junction degradation / regulation of G0 to G1 transition / Folding of actin by CCT/TriC / Cell-extracellular matrix interactions / protein localization to adherens junction / dense body / postsynaptic actin cytoskeleton / protein antigen binding / Tat protein binding / chromatin-protein adaptor activity / Ino80 complex / blastocyst formation / Prefoldin mediated transfer of substrate to CCT/TriC / RSC-type complex / regulation of double-strand break repair / regulation of nucleotide-excision repair / Adherens junctions interactions / RHOF GTPase cycle / adherens junction assembly / apical protein localization / box C/D snoRNP assembly / Sensory processing of sound by outer hair cells of the cochlea / protein folding chaperone complex / Interaction between L1 and Ankyrins / tight junction / SWI/SNF complex / regulation of mitotic metaphase/anaphase transition / Sensory processing of sound by inner hair cells of the cochlea / nucleosomal DNA binding / positive regulation of T cell differentiation / Formation of Senescence-Associated Heterochromatin Foci (SAHF) / apical junction complex / regulation of norepinephrine uptake / positive regulation of double-strand break repair / spinal cord development / transporter regulator activity / negative regulation of gene expression, epigenetic / maintenance of blood-brain barrier / regulation of chromosome organization / nitric-oxide synthase binding / cortical cytoskeleton / establishment or maintenance of cell polarity / RUNX1 interacts with co-factors whose precise effect on RUNX1 targets is not known / Transcriptional Regulation by E2F6 / NuA4 histone acetyltransferase complex / positive regulation of stem cell population maintenance / Regulation of MITF-M-dependent genes involved in pigmentation / Recycling pathway of L1 / TFIID-class transcription factor complex binding / regulation of DNA replication / brush border / regulation of G1/S transition of mitotic cell cycle / MLL1 complex / regulation of embryonic development / Telomere Extension By Telomerase / kinesin binding / EPH-ephrin mediated repulsion of cells / negative regulation of cell differentiation / somatic stem cell population maintenance / spermatid development / RHO GTPases Activate WASPs and WAVEs / regulation of synaptic vesicle endocytosis / enzyme-substrate adaptor activity / positive regulation of myoblast differentiation / RHO GTPases activate IQGAPs / RNA polymerase II core promoter sequence-specific DNA binding / heterochromatin / regulation of DNA repair / regulation of protein localization to plasma membrane / positive regulation of double-strand break repair via homologous recombination / Replacement of protamines by nucleosomes in the male pronucleus / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / EPHB-mediated forward signaling 類似検索 - 分子機能
Enhancer of polycomb, C-terminal / Enhancer of Polycomb C-terminus / DNA methyltransferase 1-associated 1 / DNA methyltransferase 1-associated protein 1 (DMAP1) / E1A-binding protein p400, N-terminal / E1A-binding protein p400, N-terminal / SWR1-complex protein 4/DNA methyltransferase 1-associated protein 1 / DAMP1, SANT/Myb-like domain / SANT/Myb-like domain of DAMP1 / Enhancer of polycomb protein ...Enhancer of polycomb, C-terminal / Enhancer of Polycomb C-terminus / DNA methyltransferase 1-associated 1 / DNA methyltransferase 1-associated protein 1 (DMAP1) / E1A-binding protein p400, N-terminal / E1A-binding protein p400, N-terminal / SWR1-complex protein 4/DNA methyltransferase 1-associated protein 1 / DAMP1, SANT/Myb-like domain / SANT/Myb-like domain of DAMP1 / Enhancer of polycomb protein / Vps72/YL1, N-terminal / YL1 nuclear protein / Vps72/YL1, C-terminal / YL1 nuclear protein C-terminal domain / YL1 nuclear protein C-terminal domain / Myb-like domain profile. / domain in helicases and associated with SANT domains / RuvB-like / RuvB-like, AAA-lid domain / RuvBL1/2, DNA/RNA binding domain / TIP49 P-loop domain / TIP49 AAA-lid domain / TIP49, P-loop domain / HSA domain / Helicase/SANT-associated domain / HSA domain profile. / Enhancer of polycomb-like, N-terminal / Enhancer of polycomb-like / : / SNF2-like, N-terminal domain superfamily / SNF2, N-terminal / SNF2-related domain / SANT/Myb domain / Actins signature 1. / Actin, conserved site / Actins signature 2. / Actin/actin-like conserved site / Actins and actin-related proteins signature. / Actin / Actin family / Actin / : / Histone H2B signature. / Histone H2A conserved site / Histone H2A signature. / Histone H2B / Histone H2B / Histone H2A, C-terminal domain / C-terminus of histone H2A / Histone 2A / Histone H2A / Helicase conserved C-terminal domain / ATPase, nucleotide binding domain / Histone H2A/H2B/H3 / Core histone H2A/H2B/H3/H4 / Histone-fold / helicase superfamily c-terminal domain / Superfamilies 1 and 2 helicase C-terminal domain profile. / Superfamilies 1 and 2 helicase ATP-binding type-1 domain profile. / DEAD-like helicases superfamily / Helicase, C-terminal / Helicase superfamily 1/2, ATP-binding domain / ATPases associated with a variety of cellular activities / AAA+ ATPase domain / P-loop containing nucleoside triphosphate hydrolase 類似検索 - ドメイン・相同性
Actin-like protein 6A / Histone H2A.Z / Histone H2B type 1-B / Actin, cytoplasmic 1 / Vacuolar protein sorting-associated protein 72 homolog / E1A-binding protein p400 / Enhancer of polycomb homolog 1 / DNA methyltransferase 1-associated protein 1 / RuvB-like 2 / RuvB-like 1 類似検索 - 構成要素
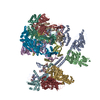
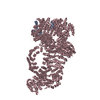
ジャーナル: Nature / 年: 2024 タイトル: Structure of the human TIP60-C histone exchange and acetyltransferase complex. 著者: Changqing Li / Ekaterina Smirnova / Charlotte Schnitzler / Corinne Crucifix / Jean Paul Concordet / Alice Brion / Arnaud Poterszman / Patrick Schultz / Gabor Papai / Adam Ben-Shem / 要旨: Chromatin structure is a key regulator of DNA transcription, replication and repair. In humans, the TIP60-EP400 complex (TIP60-C) is a 20-subunit assembly that affects chromatin structure through two ...Chromatin structure is a key regulator of DNA transcription, replication and repair. In humans, the TIP60-EP400 complex (TIP60-C) is a 20-subunit assembly that affects chromatin structure through two enzymatic activities: ATP-dependent exchange of histone H2A-H2B for H2A.Z-H2B, and histone acetylation. In yeast, however, these activities are performed by two independent complexes-SWR1 and NuA4, respectively. How the activities of the two complexes are merged into one supercomplex in humans, and what this association entails for the structure and mechanism of the proteins and their recruitment to chromatin, are unknown. Here we describe the structure of the endogenous human TIP60-C. We find a three-lobed architecture composed of SWR1-like (SWR1L) and NuA4-like (NuA4L) parts, which associate with a TRRAP activator-binding module. The huge EP400 subunit contains the ATPase motor, traverses the junction between SWR1L and NuA4L twice and constitutes the scaffold of the three-lobed architecture. NuA4L is completely rearranged compared with its yeast counterpart. TRRAP is flexibly tethered to NuA4L-in stark contrast to its robust connection to the completely opposite side of NuA4 in yeast. A modelled nucleosome bound to SWR1L, supported by tests of TIP60-C activity, suggests that some aspects of the histone exchange mechanism diverge from what is seen in yeast. Furthermore, a fixed actin module (as opposed to the mobile actin subcomplex in SWR1; ref. ), the flexibility of TRRAP and the weak effect of extranucleosomal DNA on exchange activity lead to a different, activator-based mode of enlisting TIP60-C to chromatin.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 フランス, 1件
フランス, 1件  引用
引用 構造の表示
構造の表示
 ダウンロードとリンク
ダウンロードとリンク emd_18794.png
emd_18794.png http://ftp.pdbj.org/pub/emdb/structures/EMD-18794
http://ftp.pdbj.org/pub/emdb/structures/EMD-18794









































 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)












































 試料の構成要素
試料の構成要素 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN

