 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8qr1 | ||||||
|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of the human Tip60 complex | ||||||
 Components Components |
| ||||||
 Keywords Keywords | TRANSCRIPTION / Eukaryotic transcription / Histone acetyltransferase / chromatin remodeling / Complex | ||||||
| Function / homology |  Function and homology information Function and homology informationpiccolo histone acetyltransferase complex / promoter-enhancer loop anchoring activity / telomerase RNA localization to Cajal body / sperm DNA condensation / histone chaperone activity / establishment of protein localization to chromatin / regulation of DNA strand elongation / positive regulation of telomere maintenance in response to DNA damage / positive regulation of norepinephrine uptake / R2TP complex ...piccolo histone acetyltransferase complex / promoter-enhancer loop anchoring activity / telomerase RNA localization to Cajal body / sperm DNA condensation / histone chaperone activity / establishment of protein localization to chromatin / regulation of DNA strand elongation / positive regulation of telomere maintenance in response to DNA damage / positive regulation of norepinephrine uptake / R2TP complex / dynein axonemal particle / bBAF complex / cellular response to cytochalasin B / neural retina development / Formation of the embryonic stem cell BAF (esBAF) complex / npBAF complex / brahma complex / nBAF complex / Formation of the canonical BAF (cBAF) complex / RPAP3/R2TP/prefoldin-like complex / regulation of transepithelial transport / Swr1 complex / morphogenesis of a polarized epithelium / Formation of the polybromo-BAF (pBAF) complex / Formation of neuronal progenitor and neuronal BAF (npBAF and nBAF) / structural constituent of postsynaptic actin cytoskeleton / Formation of annular gap junctions / Formation of the dystrophin-glycoprotein complex (DGC) / protein antigen binding / GBAF complex / Gap junction degradation / Formation of the non-canonical BAF (ncBAF) complex / protein localization to adherens junction / regulation of G0 to G1 transition / Cell-extracellular matrix interactions / dense body / Folding of actin by CCT/TriC / Tat protein binding / Ino80 complex / postsynaptic actin cytoskeleton / RSC-type complex / blastocyst formation / Regulation of CDH1 Function / regulation of double-strand break repair / Prefoldin mediated transfer of substrate to CCT/TriC / regulation of nucleotide-excision repair / Adherens junctions interactions / box C/D snoRNP assembly / adherens junction assembly / chromatin-protein adaptor activity / RHOF GTPase cycle / apical protein localization / Sensory processing of sound by outer hair cells of the cochlea / regulation of mitotic metaphase/anaphase transition / tight junction / SWI/SNF complex / Sensory processing of sound by inner hair cells of the cochlea / Interaction between L1 and Ankyrins / positive regulation of T cell differentiation / Formation of Senescence-Associated Heterochromatin Foci (SAHF) / apical junction complex / positive regulation of double-strand break repair / negative regulation of gene expression, epigenetic / spinal cord development / regulation of chromosome organization / maintenance of blood-brain barrier / regulation of norepinephrine uptake / transporter regulator activity / positive regulation of stem cell population maintenance / NuA4 histone acetyltransferase complex / Transcriptional Regulation by E2F6 / Recycling pathway of L1 / Regulation of MITF-M-dependent genes involved in pigmentation / cortical cytoskeleton / establishment or maintenance of cell polarity / RUNX1 interacts with co-factors whose precise effect on RUNX1 targets is not known / regulation of DNA replication / TFIID-class transcription factor complex binding / nitric-oxide synthase binding / MLL1 complex / regulation of G1/S transition of mitotic cell cycle / brush border / regulation of embryonic development / somatic stem cell population maintenance / EPH-ephrin mediated repulsion of cells / Telomere Extension By Telomerase / spermatid development / protein folding chaperone complex / negative regulation of cell differentiation / regulation of synaptic vesicle endocytosis / enzyme-substrate adaptor activity / positive regulation of myoblast differentiation / RHO GTPases Activate WASPs and WAVEs / kinesin binding / regulation of DNA repair / RNA polymerase II core promoter sequence-specific DNA binding / regulation of protein localization to plasma membrane / RHO GTPases activate IQGAPs / positive regulation of double-strand break repair via homologous recombination / EPHB-mediated forward signaling Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 2.4 Å | ||||||
 Authors Authors | Li, C. / Smirnova, E. / Schnitzler, C. / Crucifix, C. / Concordet, J.P. / Brion, A. / Poterszman, A. / Schultz, P. / Papai, G. / Ben-Shem, A. | ||||||
| Funding support |  France, 1items France, 1items
| ||||||
 Citation Citation | Journal: Nature / Year: 2024 Title: Structure of the human TIP60-C histone exchange and acetyltransferase complex. Authors: Changqing Li / Ekaterina Smirnova / Charlotte Schnitzler / Corinne Crucifix / Jean Paul Concordet / Alice Brion / Arnaud Poterszman / Patrick Schultz / Gabor Papai / Adam Ben-Shem / Abstract: Chromatin structure is a key regulator of DNA transcription, replication and repair. In humans, the TIP60-EP400 complex (TIP60-C) is a 20-subunit assembly that affects chromatin structure through two ...Chromatin structure is a key regulator of DNA transcription, replication and repair. In humans, the TIP60-EP400 complex (TIP60-C) is a 20-subunit assembly that affects chromatin structure through two enzymatic activities: ATP-dependent exchange of histone H2A-H2B for H2A.Z-H2B, and histone acetylation. In yeast, however, these activities are performed by two independent complexes-SWR1 and NuA4, respectively. How the activities of the two complexes are merged into one supercomplex in humans, and what this association entails for the structure and mechanism of the proteins and their recruitment to chromatin, are unknown. Here we describe the structure of the endogenous human TIP60-C. We find a three-lobed architecture composed of SWR1-like (SWR1L) and NuA4-like (NuA4L) parts, which associate with a TRRAP activator-binding module. The huge EP400 subunit contains the ATPase motor, traverses the junction between SWR1L and NuA4L twice and constitutes the scaffold of the three-lobed architecture. NuA4L is completely rearranged compared with its yeast counterpart. TRRAP is flexibly tethered to NuA4L-in stark contrast to its robust connection to the completely opposite side of NuA4 in yeast. A modelled nucleosome bound to SWR1L, supported by tests of TIP60-C activity, suggests that some aspects of the histone exchange mechanism diverge from what is seen in yeast. Furthermore, a fixed actin module (as opposed to the mobile actin subcomplex in SWR1; ref. ), the flexibility of TRRAP and the weak effect of extranucleosomal DNA on exchange activity lead to a different, activator-based mode of enlisting TIP60-C to chromatin. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8qr1.cif.gz | 1 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8qr1.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 8qr1.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/qr/8qr1ftp://data.pdbj.org/pub/pdb/validation_reports/qr/8qr1 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  18611MC  8qriC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |









-Links
 PDBj
PDBj



























- Assembly
Assembly
| Deposited unit | 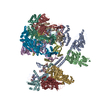
|
|---|---|
| 1 |
|
-Components
-Protein , 8 types, 13 molecules ACFSBGKLIEHDJ
| #1: Protein | Mass: 343867.312 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: K562References: UniProt: Q96L91, Hydrolases; Acting on acid anhydrides; Acting on acid anhydrides to facilitate cellular and subcellular movement | ||||||
|---|---|---|---|---|---|---|---|
| #2: Protein | Mass: 93589.172 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: K562 / References: UniProt: Q9H2F5 | ||||||
| #3: Protein | Mass: 53090.699 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: K562 / References: UniProt: Q9NPF5 | ||||||
| #4: Protein | Mass: 40658.363 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: K562 / References: UniProt: Q15906 | ||||||
| #5: Protein | Mass: 41782.660 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: K562 / References: UniProt: P60709#6: Protein | | Mass: 47509.812 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: K562 / References: UniProt: O96019#7: Protein | Mass: 50296.914 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: K562 / References: UniProt: Q9Y265, DNA helicase#8: Protein | Mass: 51222.465 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: K562 / References: UniProt: Q9Y230, DNA helicase |
-Details
| Has protein modification | N |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Tip60 complex / Type: CELL / Entity ID: all / Source: NATURAL | ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source (natural) | Organism: Homo sapiens (human) / Strain: K562 / Cellular location: nucleus | ||||||||||||||||||||||||||||||||
| Buffer solution | pH: 8 | ||||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||||
| Specimen | Conc.: 0.2 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 95 % / Chamber temperature: 279 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Microscopy | Model: FEI TITAN KRIOS | ||||||||||||
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM | ||||||||||||
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 2800 nm / Nominal defocus min: 1200 nm / Cs: 2.7 mm / Alignment procedure: COMA FREE | ||||||||||||
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER | ||||||||||||
| Image recording |
|
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 284286 | ||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 2.4 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 180397 / Symmetry type: POINT | ||||||||||||||||||||||||||||||
| Atomic model building | Protocol: RIGID BODY FIT / Space: REAL |
