 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
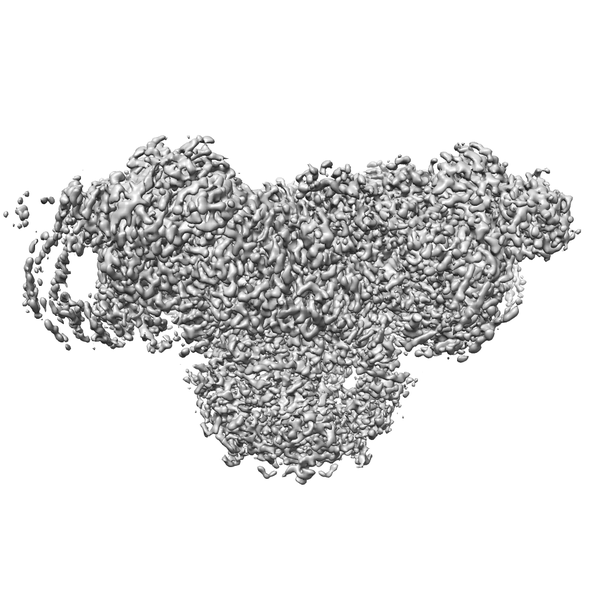
Yorodumi- EMDB-22015: Wt pig RyR1 in complex with apoCaM, EGTA condition (class 1 and 2... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: EMDB / ID: EMD-22015 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Wt pig RyR1 in complex with apoCaM, EGTA condition (class 1 and 2, closed) | |||||||||
 Map data Map data | RyR1, FKBP12.6, CaM | |||||||||
 Sample Sample |
| |||||||||
 Keywords Keywords | receptor / calcium / channel / complex / TRANSPORT PROTEIN-ISOMERASE-CALCIUM BINDING PROTEIN complex | |||||||||
| Function / homology |  Function and homology information Function and homology information: / negative regulation of calcium-mediated signaling / negative regulation of insulin secretion involved in cellular response to glucose stimulus / neuronal action potential propagation / insulin secretion involved in cellular response to glucose stimulus / negative regulation of release of sequestered calcium ion into cytosol / response to redox state / CaM pathway / Cam-PDE 1 activation / Sodium/Calcium exchangers ...: / negative regulation of calcium-mediated signaling / negative regulation of insulin secretion involved in cellular response to glucose stimulus / neuronal action potential propagation / insulin secretion involved in cellular response to glucose stimulus / negative regulation of release of sequestered calcium ion into cytosol / response to redox state / CaM pathway / Cam-PDE 1 activation / Sodium/Calcium exchangers / negative regulation of heart rate / Calmodulin induced events / 'de novo' protein folding / Reduction of cytosolic Ca++ levels / Activation of Ca-permeable Kainate Receptor / CREB1 phosphorylation through the activation of CaMKII/CaMKK/CaMKIV cascasde / Loss of phosphorylation of MECP2 at T308 / CREB1 phosphorylation through the activation of Adenylate Cyclase / negative regulation of high voltage-gated calcium channel activity / PKA activation / CaMK IV-mediated phosphorylation of CREB / FK506 binding / Glycogen breakdown (glycogenolysis) / CLEC7A (Dectin-1) induces NFAT activation / negative regulation of ryanodine-sensitive calcium-release channel activity / organelle localization by membrane tethering / Activation of RAC1 downstream of NMDARs / : / autophagosome membrane docking / negative regulation of calcium ion export across plasma membrane / regulation of ryanodine-sensitive calcium-release channel activity / regulation of cardiac muscle cell action potential / presynaptic endocytosis / Synthesis of IP3 and IP4 in the cytosol / Phase 0 - rapid depolarisation / Negative regulation of NMDA receptor-mediated neuronal transmission / Unblocking of NMDA receptors, glutamate binding and activation / RHO GTPases activate PAKs / calcineurin-mediated signaling / regulation of cell communication by electrical coupling involved in cardiac conduction / smooth muscle contraction / Ion transport by P-type ATPases / Uptake and function of anthrax toxins / protein phosphatase activator activity / Long-term potentiation / Calcineurin activates NFAT / T cell proliferation / Regulation of MECP2 expression and activity / DARPP-32 events / Smooth Muscle Contraction / detection of calcium ion / regulation of cardiac muscle contraction / catalytic complex / RHO GTPases activate IQGAPs / calcium channel inhibitor activity / presynaptic cytosol / Activation of AMPK downstream of NMDARs / cellular response to interferon-beta / regulation of release of sequestered calcium ion into cytosol by sarcoplasmic reticulum / Ion homeostasis / eNOS activation / Tetrahydrobiopterin (BH4) synthesis, recycling, salvage and regulation / Protein methylation / release of sequestered calcium ion into cytosol / titin binding / regulation of cardiac muscle contraction by regulation of the release of sequestered calcium ion / regulation of calcium-mediated signaling / voltage-gated potassium channel complex / sarcoplasmic reticulum membrane / FCERI mediated Ca+2 mobilization / calcium channel complex / substantia nigra development / FCGR3A-mediated IL10 synthesis / regulation of heart rate / Antigen activates B Cell Receptor (BCR) leading to generation of second messengers / calyx of Held / Ras activation upon Ca2+ influx through NMDA receptor / adenylate cyclase activator activity / VEGFR2 mediated cell proliferation / VEGFR2 mediated vascular permeability / protein serine/threonine kinase activator activity / regulation of cytokinesis / spindle microtubule / peptidylprolyl isomerase / positive regulation of receptor signaling pathway via JAK-STAT / sarcomere / peptidyl-prolyl cis-trans isomerase activity / Translocation of SLC2A4 (GLUT4) to the plasma membrane / calcium channel regulator activity / calcium-mediated signaling / Transcriptional activation of mitochondrial biogenesis / protein maturation / RAF activation / response to calcium ion / cellular response to type II interferon / G2/M transition of mitotic cell cycle / Stimuli-sensing channels / Z disc / spindle pole / calcium-dependent protein binding Similarity search - Function | |||||||||
| Biological species |   Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 3.8 Å | |||||||||
 Authors Authors | Woll KW / Haji-Ghassemi O | |||||||||
| Funding support | 1 items
| |||||||||
 Citation Citation | Journal: Nat Commun / Year: 2021 Title: Pathological conformations of disease mutant Ryanodine Receptors revealed by cryo-EM. Authors: Kellie A Woll / Omid Haji-Ghassemi / Filip Van Petegem /  Abstract: Ryanodine Receptors (RyRs) are massive channels that release Ca from the endoplasmic and sarcoplasmic reticulum. Hundreds of mutations are linked to malignant hyperthermia (MH), myopathies, and ...Ryanodine Receptors (RyRs) are massive channels that release Ca from the endoplasmic and sarcoplasmic reticulum. Hundreds of mutations are linked to malignant hyperthermia (MH), myopathies, and arrhythmias. Here, we explore the first MH mutation identified in humans by providing cryo-EM snapshots of the pig homolog, R615C, showing that it affects an interface between three solenoid regions. We also show the impact of apo-calmodulin (apoCaM) and how it can induce opening by bending of the bridging solenoid, mediated by its N-terminal lobe. For R615C RyR1, apoCaM binding abolishes a pathological 'intermediate' conformation, distributing the population to a mixture of open and closed channels, both different from the structure without apoCaM. Comparisons show that the mutation primarily affects the closed state, inducing partial movements linked to channel activation. This shows that disease mutations can cause distinct pathological conformations of the RyR and facilitate channel opening by disrupting interactions between different solenoid regions. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | EM map: SurfViewMolmilJmol/JSmol |
| Supplemental images |
- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_22015.map.gz | 62.8 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-22015-v30.xmlemd-22015.xml | 18.7 KB 18.7 KB | Display Display | EMDB header |
| Images | 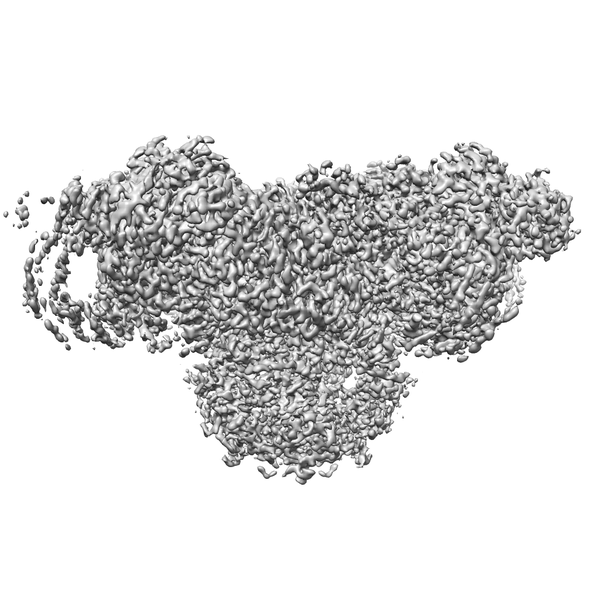 emd_22015.png emd_22015.png | 222.6 KB | ||
| Filedesc metadata | emd-22015.cif.gz | 7.9 KB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-22015ftp://ftp.pdbj.org/pub/emdb/structures/EMD-22015 http://ftp.pdbj.org/pub/emdb/structures/EMD-22015ftp://ftp.pdbj.org/pub/emdb/structures/EMD-22015 | HTTPS FTP |
-Related structure data
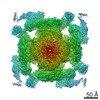
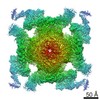
| Related structure data |  6x32MC  6w1nC  6x33C  6x34C  6x35C  6x36C C: citing same article ( M: atomic model generated by this map |
|---|---|
| Similar structure data |









-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| Related items in Molecule of the Month |



















-Map
| File | Download / File: emd_22015.map.gz / Format: CCP4 / Size: 421.9 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annotation | RyR1, FKBP12.6, CaM | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 1.09 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Density |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
CCP4 map header:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-Supplemental data
- Sample components
Sample components
-Entire : ryanodine receptor-FKBP1B-Calmodulin complex
| Entire | Name: ryanodine receptor-FKBP1B-Calmodulin complex |
|---|---|
| Components |
|
-Supramolecule #1: ryanodine receptor-FKBP1B-Calmodulin complex
| Supramolecule | Name: ryanodine receptor-FKBP1B-Calmodulin complex / type: complex / ID: 1 / Parent: 0 / Macromolecule list: #1-#3 |
|---|
-Supramolecule #2: ryanodine receptor
| Supramolecule | Name: ryanodine receptor / type: complex / ID: 2 / Parent: 1 / Macromolecule list: #2 |
|---|---|
| Source (natural) | Organism: |
-Supramolecule #3: FKBP1B
| Supramolecule | Name: FKBP1B / type: complex / ID: 3 / Parent: 1 / Macromolecule list: #1 |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
-Supramolecule #4: Calmodulin
| Supramolecule | Name: Calmodulin / type: complex / ID: 4 / Parent: 1 / Macromolecule list: #3 |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
-Macromolecule #1: Peptidyl-prolyl cis-trans isomerase FKBP1B
| Macromolecule | Name: Peptidyl-prolyl cis-trans isomerase FKBP1B / type: protein_or_peptide / ID: 1 / Number of copies: 4 / Enantiomer: LEVO / EC number: peptidylprolyl isomerase |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 11.939562 KDa |
| Recombinant expression | Organism:  |
| Sequence | String: SNAGVEIETI SPGDGRTFPK KGQTCVVHYT GMLQNGKKFD SSRDRNKPFK FRIGKQEVIK GFEEGAAQMS LGQRAKLTCT PDVAYGATG HPGVIPPNAT LIFDVELLNL E UniProtKB: Peptidyl-prolyl cis-trans isomerase FKBP1B |
-Macromolecule #2: Ryanodine Receptor
| Macromolecule | Name: Ryanodine Receptor / type: protein_or_peptide / ID: 2 / Number of copies: 4 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: |
| Molecular weight | Theoretical: 422.853312 KDa |
| Sequence | String: QFLRTDDEVV LQCNATVLKE QLKLCLAAEG FGNRLCFLEP TSNAQNVPPD LAICCFVLEQ SLSVRALQEM LANTVEAGHR TLLYGHAIL LRHAHSGMYL SCLTTSRSMT DKLAFDVGLQ EDATGEACWW TTHPASKQRS EGEKVRVGDD LILVSVSSER Y LHLSTLQV ...String: QFLRTDDEVV LQCNATVLKE QLKLCLAAEG FGNRLCFLEP TSNAQNVPPD LAICCFVLEQ SLSVRALQEM LANTVEAGHR TLLYGHAIL LRHAHSGMYL SCLTTSRSMT DKLAFDVGLQ EDATGEACWW TTHPASKQRS EGEKVRVGDD LILVSVSSER Y LHLSTLQV DASFMQTLWN MNPICSGCEE GYVTGGHVLR LFHGHMDECL TISPADSDDQ RRLVYYEGGS VCTHARSLWR LE PLRISWS GSHLRWGQPL RIRHVTTGRY LALIEDQGLV VVDASKAHTK ATSFCFRISK EKLKRDVEGM GPPEIKYGES LCF VQHVAS GLWLTYAALK KKAILHQEGH MDDALSLTRC QQEESQAARM IYSTAGLYNH FIKGLDSFSG KPRGSGAPAG TALP LEGVI LSLQDLIGYF EPPSEELQHE EKQSKLRSLR NRQSLFQEEG MLSLVLNCID RLNVYTTAAH FAEFAGEEAA ESWKE IVNL LYEILASLIR GNRANCALFS NNLDWLVSKL DRLEASSGIL EVLYCVLIES PEVLNIIQEN HIKSIISLLD KHGRNH KVL DVLCSLCVCN GVAVRSNQDL ITENLLPGRE LLLQTNLINY VTSIRPNIFV GRAEGTTQYS KWYFEVMVDE VVPFLTA QA THLRVGWALT EGYSPYPGGG EGWGGNGVGD DLYSYGFDGL HLWTGHVPRL VTSPGQHLLA PEDVVSCCLD LSVPSISF R INGCPVQGVF EAFNLNGLFF PVVSFSAGVK VRFLLGGRHG EFKFLPPPGY APCHEAVLPR ERLRLEPIKE YRREGPRGP HLVGPSRCLS HTDFVPCPLP PHLERIREKL AENIHELWAL TRIEQGWTYG PVRDDNKRLH PCLVDFHSLP EPERNYNLQM SGETLKTLL ALGCHVGMAD EKAEDNLRKT KLPKTYMMSN GYKPAPLDLS HVRLTPAQTT LVDRLAENGH NVWARDRVAQ G WSYSAVQD IPARRNPRLV PYRLLDEATK RSNRDSLCQA VRTLLGYGYN IERVRIFRAE KSYAVQSGRW YFEFEAVTTG EM RVGWARP ELRPDVELGA DELAYVFNGH RGQRWHLGSE LFGRPWQSGD VVGCMIDLTE NTIIFTLNGE VLMSDSGSET AFR DIEVGD GFLPVCSLGP GQVGHLNLGQ DVSSLRFFAI CGLQEGFEPF AINMQRPVTT WFSKSLPQFE AVPLEHPHYE VSRV DGTVD TPPCLRLTHR SLVEMLFLRL SLPVQFHQLN TTTYYYSVRV FAGQEPSCVW VGWVTPDYHQ HDMNFDLTKV RAVTV TMGD NIHSSLKCSN CYMVWGGDFV SHTDLVIGCL VDLATGLMTF TANGKESNTF FQVEPNTKLF PAVFVLPTHQ NVIQFE LGK QKNIMPLSAA MFLSERKNPA PQCPPRLEMQ MLMPVSWSRM PNHFLRVETR RAGERLGWAV QCQEPLTMMA LHIPEEN RC MDILELSERL DLQQFHSHTL RLYRAVCALG NNRVAHALCS HVDQAQLLHA LEDAHLPGPL RAGYYDLLIS IHLESACR S RRSMLSEYIV PLTPETRAIT LFPPRRHGLP GVGVTTSLRP PHHFSAPCFV AALPEAPARL SPSIPLEALR DKALRMLGE AVRDGGQHAR DPVGGSVEFQ FVPVLKLVST LLVMGIFGDE DVKQILKMIE PEVEEGLLQM KLPESVKLQM CNLLEYFCDQ ELQHRVESL AAFAERYVDK LQANQRDRYG ILMKAFTMTA AETARRTREF RSPPQEQINM LLHFKPLPDE IRQDLLEFHQ D LLTHCGIQ LQSLQELVSH TVVRWAQEDF VQSPELVRAM FSLLHRQYDG LGELLRALPR AYTISPSSVE DTMSLLECLG QI RSLLIVQ MGPQEENLMI QSIGNIMNNK VFYQHPNLMR ALGMHETVME VMVNVLGKEI RFPKMVTSCC RFLCYFCRIS RQN QRSMFD HLSYLLENSG IGLGMQGSTP LDVAAASVID NNELALALQE QDLEKVVSYL AGCGLQSYPD IGWNPCGGER YLDF LRFAV FVNGESVEEN ANVVVRLLIR KPECFGPALR GEGGSGLLAT IEEAIRISEH LGHAIMSFYA ALIDLLGRCA PEMHL IQAG KGEALRIRAI LRSLVPLDDL VGIISLPKMS ASFVPDHKAS MVLFLDRVYG IENAAFLLHV LDVGFLPDMR AAATFS TTE MALALNRYLC LAVLPLITKC APLFAMVDSM LHTVYRLSRG RSLTKAQRDV IEECLMALCR YIRPSMLQHL LRRLVFD VP (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)AADPRPVET LNVIIPEKLD SFINKFAEYT HEKWAFDKIQ NNWSYGENID EELKTHPMLR PYKTFSEKDK EIYRWP IKE SLKAMIAWEW TIEKAREGEY NPQPPDLSGV TLSRELQAMA EQLAENYHNT WGRKKKQELE AKGGGTHPLL VPYDTLT AK EKARDREKAQ ELLKFLQMNG YAVTR(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)PLLIRY VDNN RAHWL TEPNPSAEEL FRMVGEIFIY WSK(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)RRAVVAC FRMTPLYNLP THRACNMFLE SYKAAWILTE DHS FEDRMI DDLSKAGEQE EEEEEVEEKK PDPLHQLVLH FSRTALTEKS KLDEDYLYMA YADIMAKSCH LSFEEKEMEK QRLL YQQAR LHNRGAAEMV LQMISACKGE TGAMVSSTLK LGISILNGGN ADVQQKMLDY LKDKKEVGFF QSIQALMQTC SVLDL NAFE RQNKAEGLGM VNEDGTVEKV MADDEFTQDL FRFLQLLCEG HNNDFQNYLR TQTGNTTTIN IIICTVDYLL RLQESI SDF YWYYSGKDVI EEQGKRNFSK AMSVAKQVFN SLTEYIQGPC TGNQQSLAHS RLWDAVVGFL HVFAHMMMKL AQDSSQI EL LKELLDLQKD MVVMLLSLLE GNVVNGMIAR QMVDMLVESS SNVEMILKFF DMFLKLKDIV GSEAFQDYVT DPRGLISK K DFQK(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) EFANRFQEPA RDIGFNVAVL LTNLSEHVPH DPRLRNFLEL AESIL EYFR PYLGRIEIMG ASRRIERIYF EISETNRAQW EMPQVKESKR QFIFDVVNEG GESEKMELFV SFCEDTIFEM QIAAQI (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)EVQRV KFLNYLSRNF YTLRFLALFL AFAINFILLF YKVSDSPPVY YFLEESTGYM EPALRCLSLL HTLVAFLCII GYNCLKVPL VIFKREKELA RKLEFDGLYI TEQPEDDDVK GQWDRLVLNT PSFPSNYWDK FVKRKVLDKH GDIYGRERIA E IDVKYQIW KFGVIFTDNS FLYLGWYMVM SLLGHYNNFF FAAHLLDIAM GVKTLRTILS SVTHNGKQLV MTVGLLAVVV YL YTVVAFN FFRKFYNKDM KCDDMMTCYL FHMYVGVRAG GGIGDEIEDP AGDEYELYRV VFDITFFFFV IVILLAIIQG LII DAFGEL RDQQEQVRED METKCFICGI GSDYFDTTPH RFETHTLEEH NLANYMFFLM YLINKDETEH TGQESYVWKM YQER CWDFF PAGDCFRKQY EDQLS |
-Macromolecule #3: Calmodulin-1
| Macromolecule | Name: Calmodulin-1 / type: protein_or_peptide / ID: 3 / Number of copies: 4 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 16.521094 KDa |
| Recombinant expression | Organism: |
| Sequence | String: DQLTEEQIAE FKEAFSLFDK DGDGTITTKE LGTVMRSLGQ NPTEAELQDM INEVDADGNG TIDFPEFLTM MARKMKDTDS EEEIREAFR VFDKDGNGYI SAAELRHVMT NLGEKLTDEE VDEMIREADI DGDGQVNYEE FVQMMTA UniProtKB: Calmodulin-1 |
-Macromolecule #4: ZINC ION
| Macromolecule | Name: ZINC ION / type: ligand / ID: 4 / Number of copies: 4 / Formula: ZN |
|---|---|
| Molecular weight | Theoretical: 65.409 Da |
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Buffer | pH: 7.5 |
|---|---|
| Grid | Details: unspecified |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy
Electron microscopy
| Microscope | FEI TITAN KRIOS |
|---|---|
| Image recording | Film or detector model: FEI FALCON III (4k x 4k) / Average electron dose: 50.0 e/Å2 |
| Electron beam | Acceleration voltage: 300 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: OTHER / Imaging mode: DIFFRACTION |
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
-Image processing
| Startup model | Type of model: PDB ENTRY |
|---|---|
| Final reconstruction | Applied symmetry - Point group: C4 (4 fold cyclic) / Resolution.type: BY AUTHOR / Resolution: 3.8 Å / Resolution method: FSC 0.143 CUT-OFF / Software - Name: PHENIX (ver. dev-3714) / Number images used: 44957 |
| Initial angle assignment | Type: MAXIMUM LIKELIHOOD |
| Final angle assignment | Type: MAXIMUM LIKELIHOOD |
