 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-6odm: Herpes simplex virus type 1 (HSV-1) portal vertex-adjacent capsid... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6odm | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Herpes simplex virus type 1 (HSV-1) portal vertex-adjacent capsid/CATC, asymmetric unit | ||||||||||||||||||||||||||||||
 Components Components |
| ||||||||||||||||||||||||||||||
 Keywords Keywords | VIRAL PROTEIN / tegument / portal / DNA-packaging / capsid | ||||||||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationT=16 icosahedral viral capsid / viral genome packaging / viral tegument / deNEDDylase activity / viral capsid assembly / viral DNA genome replication / viral process / chromosome organization / virion component / viral penetration into host nucleus ...T=16 icosahedral viral capsid / viral genome packaging / viral tegument / deNEDDylase activity / viral capsid assembly / viral DNA genome replication / viral process / chromosome organization / virion component / viral penetration into host nucleus / viral capsid / host cell / host cell cytoplasm / symbiont-mediated perturbation of host ubiquitin-like protein modification / cysteine-type deubiquitinase activity / symbiont entry into host cell / host cell nucleus / structural molecule activity / proteolysis / DNA binding Similarity search - Function | ||||||||||||||||||||||||||||||
| Biological species |   Human herpesvirus 1 strain KOS Human herpesvirus 1 strain KOS | ||||||||||||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4.3 Å | ||||||||||||||||||||||||||||||
 Authors Authors | Liu, Y.T. / Jih, J. / Dai, X.H. / Bi, G.Q. / Zhou, Z.H. | ||||||||||||||||||||||||||||||
| Funding support |  China, China,  United States, 9items United States, 9items
| ||||||||||||||||||||||||||||||
 Citation Citation | Journal: Nature / Year: 2019 Title: Cryo-EM structures of herpes simplex virus type 1 portal vertex and packaged genome. Authors: Yun-Tao Liu / Jonathan Jih / Xinghong Dai / Guo-Qiang Bi / Z Hong Zhou / Abstract: Herpesviruses are enveloped viruses that are prevalent in the human population and are responsible for diverse pathologies, including cold sores, birth defects and cancers. They are characterized by ...Herpesviruses are enveloped viruses that are prevalent in the human population and are responsible for diverse pathologies, including cold sores, birth defects and cancers. They are characterized by a highly pressurized pseudo-icosahedral capsid-with triangulation number (T) equal to 16-encapsidating a tightly packed double-stranded DNA (dsDNA) genome. A key process in the herpesvirus life cycle involves the recruitment of an ATP-driven terminase to a unique portal vertex to recognize, package and cleave concatemeric dsDNA, ultimately giving rise to a pressurized, genome-containing virion. Although this process has been studied in dsDNA phages-with which herpesviruses bear some similarities-a lack of high-resolution in situ structures of genome-packaging machinery has prevented the elucidation of how these multi-step reactions, which require close coordination among multiple actors, occur in an integrated environment. To better define the structural basis of genome packaging and organization in herpes simplex virus type 1 (HSV-1), we developed sequential localized classification and symmetry relaxation methods to process cryo-electron microscopy (cryo-EM) images of HSV-1 virions, which enabled us to decouple and reconstruct hetero-symmetric and asymmetric elements within the pseudo-icosahedral capsid. Here we present in situ structures of the unique portal vertex, genomic termini and ordered dsDNA coils in the capsid spooled around a disordered dsDNA core. We identify tentacle-like helices and a globular complex capping the portal vertex that is not observed in phages, indicative of herpesvirus-specific adaptations in the DNA-packaging process. Finally, our atomic models of portal vertex elements reveal how the fivefold-related capsid accommodates symmetry mismatch imparted by the dodecameric portal-a longstanding mystery in icosahedral viruses-and inform possible DNA-sequence recognition and headful-sensing pathways involved in genome packaging. This work showcases how to resolve symmetry-mismatched elements in a large eukaryotic virus and provides insights into the mechanisms of herpesvirus genome packaging. | ||||||||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6odm.cif.gz | 1.6 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6odm.ent.gz | 1.2 MB | Display | PDB format |
| PDBx/mmJSON format | 6odm.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/od/6odmftp://data.pdbj.org/pub/pdb/validation_reports/od/6odm | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  9860MC 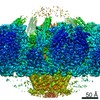 9861C 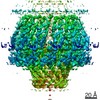 9862C  9863C 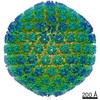 9864C 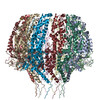 6od7C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |



-Links
 PDBj
PDBj
- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 3 types, 10 molecules TWSXLPEFOB
| #1: Protein | Mass: 149303.125 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Details: P2 MCP subunit of periportal P hexon / Source: (natural) Human herpesvirus 1 strain KOS / Strain: KOS / References: UniProt: H9E925#2: Protein | Mass: 12108.655 Da / Num. of mol.: 4 / Source method: isolated from a natural source Details: Chain E: P1 SCP subunit of periportal P hexon; decorates periportal P1 MCP (chain S) Chain F: P2 SCP subunit of periportal P hexon; decorates periportal P2 MCP (chain T) Chain L: P5 SCP ...Details: Chain E: P1 SCP subunit of periportal P hexon; decorates periportal P1 MCP (chain S) Chain F: P2 SCP subunit of periportal P hexon; decorates periportal P2 MCP (chain T) Chain L: P5 SCP subunit of periportal P hexon; decorates periportal P5 MCP (chain W) Chain P: P6 SCP subunit of periportal P hexon; decorates periportal P6 MCP (chain X) Source: (natural) Human herpesvirus 1 strain KOS / Strain: KOS / References: UniProt: Q25BW6#7: Protein | Mass: 329179.500 Da / Num. of mol.: 2 / Source method: isolated from a natural source Details: Chain B: left pUL36 subunit of the periportal capsid-associated tegument complex (CATC); each CATC contains two pUL36 subunits Chain O: right pUL36 subunit of the periportal CATC Source: (natural) Human herpesvirus 1 strain KOS / Strain: KOSReferences: UniProt: A0A0B5E3K2, UniProt: I1UYK0*PLUS, ubiquitinyl hydrolase 1, Hydrolases; Acting on peptide bonds (peptidases); Cysteine endopeptidases |
|---|
-Triplex capsid protein ... , 2 types, 6 molecules 5D67HA
| #3: Protein | Mass: 50370.363 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Details: Tri1 subunit of periportal triplex Ta / Source: (natural) Human herpesvirus 1 strain KOS / Strain: KOS / References: UniProt: Q1T724#4: Protein | Mass: 34301.617 Da / Num. of mol.: 4 / Source method: isolated from a natural source Details: Chain 6: Tri2B subunit of periportal triplex Ta Chain H: Tri2B subunit of periportal triplex Tc Source: (natural) Human herpesvirus 1 strain KOS / Strain: KOS / References: UniProt: G8H8D9 |
|---|
-Capsid vertex component ... , 2 types, 3 molecules CGK
| #5: Protein | Mass: 74769.391 Da / Num. of mol.: 1 / Source method: isolated from a natural source Details: pUL17 subunit of the periportal capsid-associated tegument complex (CATC); each CATC contains one pUL17 subunit Source: (natural) Human herpesvirus 1 strain KOS / Strain: KOS / References: UniProt: F8REV0 |
|---|---|
| #6: Protein | Mass: 62752.777 Da / Num. of mol.: 2 / Source method: isolated from a natural source Details: upper pUL25 subunit of the periportal capsid-associated tegument complex (CATC); each CATC contains two pUL25 subunits Source: (natural) Human herpesvirus 1 strain KOS / Strain: KOS / References: UniProt: D3YPI2 |
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Human herpesvirus 1 strain KOS / Type: VIRUS / Details: Cultured in Vero cells. / Entity ID: all / Source: NATURAL |
|---|---|
| Molecular weight | Value: 200 MDa / Experimental value: NO |
| Source (natural) | Organism: Human herpesvirus 1 strain KOS |
| Details of virus | Empty: NO / Enveloped: YES / Isolate: STRAIN / Type: VIRION |
| Natural host | Organism: Homo sapiens |
| Virus shell | Name: Capsid / Diameter: 1300 nm / Triangulation number (T number): 16 |
| Buffer solution | pH: 7.4 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Grid material: COPPER / Grid type: Quantifoil R2/1 |
| Vitrification | Instrument: HOMEMADE PLUNGER / Cryogen name: ETHANE / Chamber temperature: 298 K Details: The sample was manually blotted and frozen with a homemade plunger. |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 14000 X / Calibrated magnification: 24271 X / Calibrated defocus min: 1000 nm / Calibrated defocus max: 3000 nm / Cs: 2.7 mm |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER / Temperature (max): 79 K |
| Image recording | Average exposure time: 13 sec. / Electron dose: 25 e/Å2 / Detector mode: SUPER-RESOLUTION / Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Num. of real images: 8000 |
| Image scans | Sampling size: 2.5 µm / Width: 1440 / Height: 1440 / Movie frames/image: 26 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 45445 | ||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C5 (5 fold cyclic) | ||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4.3 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 42857 / Algorithm: FOURIER SPACE / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||
| Atomic model building | B value: 100 / Protocol: FLEXIBLE FIT / Space: REAL Details: Capsid and CATC atomic models from Dai & Zhou, Science 2018 (PDB-6CGR) were docked into the C5 portal vertex map (EMD-9860) and manually refined in Coot to improve fit. Regions demonstrating ...Details: Capsid and CATC atomic models from Dai & Zhou, Science 2018 (PDB-6CGR) were docked into the C5 portal vertex map (EMD-9860) and manually refined in Coot to improve fit. Regions demonstrating major differences were reconstructed ab initio. Models were then iteratively refined between real space refinement in PHENIX and manual refinement in Coot. | ||||||||||||||||||||||||||||||||||||||||
| Refinement | Stereochemistry target values: GeoStd + Monomer Library | ||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|
