ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6nzd | ||||||
|---|---|---|---|---|---|---|---|
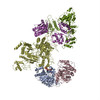
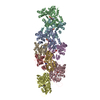
| タイトル | Cryo-EM Structure of the Lysosomal Folliculin Complex (FLCN-FNIP2-RagA-RagC-Ragulator) | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | signaling protein/inhibitor / Lysosome / mTORC1 regulation / Amino acid sensing / GTPase / SIGNALING PROTEIN / signaling protein-inhibitor complex | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報negative regulation of cell proliferation involved in kidney development / cell proliferation involved in kidney development / negative regulation of post-translational protein modification / regulation of cholesterol import / positive regulation of protein localization to lysosome / regulation of cell-substrate junction organization / Gtr1-Gtr2 GTPase complex / regulation of cholesterol efflux / positive regulation of RNA polymerase II regulatory region sequence-specific DNA binding / FNIP-folliculin RagC/D GAP ...negative regulation of cell proliferation involved in kidney development / cell proliferation involved in kidney development / negative regulation of post-translational protein modification / regulation of cholesterol import / positive regulation of protein localization to lysosome / regulation of cell-substrate junction organization / Gtr1-Gtr2 GTPase complex / regulation of cholesterol efflux / positive regulation of RNA polymerase II regulatory region sequence-specific DNA binding / FNIP-folliculin RagC/D GAP / Ragulator complex / negative regulation of brown fat cell differentiation / regulation of Ras protein signal transduction / protein localization to cell junction / regulation of TORC1 signaling / regulation of pro-B cell differentiation / negative regulation of lysosome organization / protein localization to lysosome / regulation of TOR signaling / TORC1 signaling / endosome organization / fibroblast migration / lysosome localization / Amino acids regulate mTORC1 / ATPase inhibitor activity / MTOR signalling / Energy dependent regulation of mTOR by LKB1-AMPK / protein localization to membrane / kinase activator activity / negative regulation of TOR signaling / enzyme inhibitor activity / negative regulation of glycolytic process / cell-cell junction assembly / enzyme-substrate adaptor activity / negative regulation of cold-induced thermogenesis / negative regulation of Rho protein signal transduction / azurophil granule membrane / endosomal transport / small GTPase-mediated signal transduction / regulation of cell size / lysosome organization / Macroautophagy / positive regulation of transforming growth factor beta receptor signaling pathway / RHOJ GTPase cycle / RHOQ GTPase cycle / mTORC1-mediated signalling / tertiary granule membrane / hemopoiesis / CDC42 GTPase cycle / ficolin-1-rich granule membrane / RHOH GTPase cycle / centriolar satellite / negative regulation of phosphatidylinositol 3-kinase/protein kinase B signal transduction / RHOG GTPase cycle / positive regulation of TOR signaling / regulation of receptor recycling / response to amino acid / RAC2 GTPase cycle / TOR signaling / RAC3 GTPase cycle / cellular response to nutrient levels / specific granule membrane / positive regulation of intrinsic apoptotic signaling pathway / positive regulation of autophagy / protein-membrane adaptor activity / energy homeostasis / tumor necrosis factor-mediated signaling pathway / RAC1 GTPase cycle / positive regulation of TORC1 signaling / intrinsic apoptotic signaling pathway / cellular response to amino acid starvation / ERK1 and ERK2 cascade / cellular response to starvation / phosphatidylinositol 3-kinase/protein kinase B signal transduction / GTPase activator activity / negative regulation of autophagy / viral genome replication / RNA splicing / transforming growth factor beta receptor signaling pathway / : / epithelial cell proliferation / guanyl-nucleotide exchange factor activity / cholesterol homeostasis / Regulation of PTEN gene transcription / positive regulation of interleukin-8 production / regulation of cell growth / cellular response to amino acid stimulus / TP53 Regulates Metabolic Genes / phosphoprotein binding / positive regulation of protein-containing complex assembly / regulation of protein phosphorylation / MAP2K and MAPK activation / response to virus / cilium / mitotic spindle / negative regulation of ERK1 and ERK2 cascade / positive regulation of protein localization to nucleus / intrinsic apoptotic signaling pathway in response to DNA damage / negative regulation of epithelial cell proliferation / GDP binding 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.6 Å | ||||||
 データ登録者 データ登録者 | Fromm, S.A. / Young, L.N. / Hurley, J.H. | ||||||
| 資金援助 |  米国, 1件 米国, 1件
| ||||||
 引用 引用 | ジャーナル: Science / 年: 2019 タイトル: Structural mechanism of a Rag GTPase activation checkpoint by the lysosomal folliculin complex. 著者: Rosalie E Lawrence / Simon A Fromm / Yangxue Fu / Adam L Yokom / Do Jin Kim / Ashley M Thelen / Lindsey N Young / Chun-Yan Lim / Avi J Samelson / James H Hurley / Roberto Zoncu / 要旨: The tumor suppressor folliculin (FLCN) enables nutrient-dependent activation of the mechanistic target of rapamycin complex 1 (mTORC1) protein kinase via its guanosine triphosphatase (GTPase) ...The tumor suppressor folliculin (FLCN) enables nutrient-dependent activation of the mechanistic target of rapamycin complex 1 (mTORC1) protein kinase via its guanosine triphosphatase (GTPase) activating protein (GAP) activity toward the GTPase RagC. Concomitant with mTORC1 inactivation by starvation, FLCN relocalizes from the cytosol to lysosomes. To determine the lysosomal function of FLCN, we reconstituted the human lysosomal FLCN complex (LFC) containing FLCN, its partner FLCN-interacting protein 2 (FNIP2), and the RagA:RagC GTPases as they exist in the starved state with their lysosomal anchor Ragulator complex and determined its cryo-electron microscopy structure to 3.6 angstroms. The RagC-GAP activity of FLCN was inhibited within the LFC, owing to displacement of a catalytically required arginine in FLCN from the RagC nucleotide. Disassembly of the LFC and release of the RagC-GAP activity of FLCN enabled mTORC1-dependent regulation of the master regulator of lysosomal biogenesis, transcription factor E3, implicating the LFC as a checkpoint in mTORC1 signaling. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6nzd.cif.gz | 350.7 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6nzd.ent.gz | 265.2 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6nzd.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 6nzd_validation.pdf.gz | 901.4 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 6nzd_full_validation.pdf.gz | 925.9 KB | 表示 | |
| XML形式データ | 6nzd_validation.xml.gz | 60.8 KB | 表示 | |
| CIF形式データ | 6nzd_validation.cif.gz | 91.8 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/nz/6nzdftp://data.pdbj.org/pub/pdb/validation_reports/nz/6nzd | HTTPS FTP |
-関連構造データ






-リンク
 PDBj
PDBj





- 集合体
集合体
| 登録構造単位 | 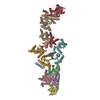
|
|---|---|
| 1 |
|
-要素
-Ragulator complex protein ... , 4種, 4分子 ABCD
| #1: タンパク質 | 分子量: 18325.350 Da / 分子数: 1 / 変異: G2A / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LAMTOR1, C11orf59, PDRO, PP7157発現宿主:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q6IAA8 |
|---|---|
| #2: タンパク質 | 分子量: 13645.579 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LAMTOR2, MAPBPIP, ROBLD3, HSPC003発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9Y2Q5 |
| #3: タンパク質 | 分子量: 13637.678 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LAMTOR3, MAP2K1IP1, MAPKSP1, PRO2783発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9UHA4 |
| #4: タンパク質 | 分子量: 10753.236 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LAMTOR4, C7orf59発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q0VGL1 |
-タンパク質 , 3種, 3分子 EHI
| #5: タンパク質 | 分子量: 18178.520 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LAMTOR5, HBXIP, hCG_40252発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: A0A0C4DGV4, UniProt: O43504*PLUS |
|---|---|
| #8: タンパク質 | 分子量: 69143.070 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: FLCN, BHD / 細胞株 (発現宿主): HEK 293 GNTI / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q8NFG4 |
| #9: タンパク質 | 分子量: 122475.414 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: FNIP2, FNIPL, KIAA1450, MAPO1 / 細胞株 (発現宿主): HEK 293 GNTI / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q9P278 |
-Ras-related GTP-binding protein ... , 2種, 2分子 FG
| #6: タンパク質 | 分子量: 36615.168 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RRAGA発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q7L523 |
|---|---|
| #7: タンパク質 | 分子量: 44758.336 Da / 分子数: 1 / 変異: D181N / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RRAGC発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9HB90 |
-非ポリマー , 2種, 2分子 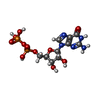

| #10: 化合物 | ChemComp-GDP /  タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM |
|---|---|
| #11: 化合物 | ChemComp-L8S /  分子量: 540.231 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N4O14P3S 分子量: 540.231 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N4O14P3S |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: FLCN-FNIP2-RagA-RagC-Ragulator Complex / タイプ: COMPLEX / 詳細: RagA bound to GDP; RagC bound to XTPgammaS / Entity ID: #1-#9 / 由来: MULTIPLE SOURCES | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.34 MDa / 実験値: NO | ||||||||||||||||||||||||||||||
| 由来(天然) | 生物種: Homo sapiens (ヒト) | ||||||||||||||||||||||||||||||
| 由来(組換発現) | 生物種: Spodoptera frugiperda (ツマジロクサヨトウ) | ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.4 | ||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.5 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 試料支持 | 詳細: 15 W / グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: C-flat-2/1 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 90 % / 凍結前の試料温度: 277 K / 詳細: Whatman 597 |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 3000 nm / 最小 デフォーカス(公称値): 1000 nm / Cs: 2.7 mm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 11 sec. / 電子線照射量: 65.6 e/Å2 / 検出モード: SUPER-RESOLUTION フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 1 / 実像数: 2703 |
| 電子光学装置 | エネルギーフィルタースリット幅: 20 eV |
| 画像スキャン | 動画フレーム数/画像: 44 |
- 解析
解析
| EMソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 982343 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.6 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 163376 / 対称性のタイプ: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | 空間: REAL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 最高解像度: 3.6 Å |