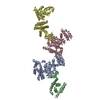
- EMDB-0554: Cryo-EM Structure of the Lysosomal Folliculin Complex (FLCN-FNIP2... -
+
データを開く
IDまたはキーワード:
読み込み中...
-
基本情報
登録情報
データベース: EMDB / ID: EMD-0554
タイトル
Cryo-EM Structure of the Lysosomal Folliculin Complex (FLCN-FNIP2-RagA-RagC-Ragulator)
マップデータ
generated by non-uniform refinement from cryoSPARC v2 followed by LocScale amplitude scaling
試料
複合体: FLCN-FNIP2-RagA-RagC-Ragulator Complex
タンパク質・ペプチド: x 9種
リガンド: x 2種
機能・相同性
機能・相同性情報
negative regulation of cell proliferation involved in kidney development / negative regulation of post-translational protein modification / cell proliferation involved in kidney development / regulation of cholesterol import / positive regulation of protein localization to lysosome / regulation of cell-substrate junction organization / Gtr1-Gtr2 GTPase complex / regulation of cholesterol efflux / positive regulation of RNA polymerase II regulatory region sequence-specific DNA binding / FNIP-folliculin RagC/D GAP ...negative regulation of cell proliferation involved in kidney development / negative regulation of post-translational protein modification / cell proliferation involved in kidney development / regulation of cholesterol import / positive regulation of protein localization to lysosome / regulation of cell-substrate junction organization / Gtr1-Gtr2 GTPase complex / regulation of cholesterol efflux / positive regulation of RNA polymerase II regulatory region sequence-specific DNA binding / FNIP-folliculin RagC/D GAP / Ragulator complex / negative regulation of brown fat cell differentiation / regulation of Ras protein signal transduction / protein localization to cell junction / regulation of TORC1 signaling / negative regulation of lysosome organization / regulation of pro-B cell differentiation / protein localization to lysosome / TORC1 signaling / regulation of TOR signaling / endosome organization / ATPase inhibitor activity / MTOR signalling / Amino acids regulate mTORC1 / fibroblast migration / lysosome localization / Energy dependent regulation of mTOR by LKB1-AMPK / protein localization to membrane / kinase activator activity / enzyme-substrate adaptor activity / negative regulation of glycolytic process / enzyme inhibitor activity / negative regulation of TOR signaling / cell-cell junction assembly / negative regulation of cold-induced thermogenesis / negative regulation of Rho protein signal transduction / azurophil granule membrane / endosomal transport / regulation of cell size / Macroautophagy / small GTPase-mediated signal transduction / lysosome organization / positive regulation of transforming growth factor beta receptor signaling pathway / RHOJ GTPase cycle / RHOQ GTPase cycle / mTORC1-mediated signalling / cellular response to nutrient levels / tertiary granule membrane / CDC42 GTPase cycle / RHOH GTPase cycle / hemopoiesis / ficolin-1-rich granule membrane / centriolar satellite / RHOG GTPase cycle / positive regulation of TOR signaling / regulation of receptor recycling / TOR signaling / RAC2 GTPase cycle / RAC3 GTPase cycle / negative regulation of phosphatidylinositol 3-kinase/protein kinase B signal transduction / response to amino acid / positive regulation of autophagy / specific granule membrane / energy homeostasis / positive regulation of intrinsic apoptotic signaling pathway / protein-membrane adaptor activity / positive regulation of TORC1 signaling / tumor necrosis factor-mediated signaling pathway / RAC1 GTPase cycle / ERK1 and ERK2 cascade / cellular response to amino acid starvation / cellular response to starvation / negative regulation of autophagy / viral genome replication / intrinsic apoptotic signaling pathway / RNA splicing / phosphatidylinositol 3-kinase/protein kinase B signal transduction / GTPase activator activity / transforming growth factor beta receptor signaling pathway / guanyl-nucleotide exchange factor activity / epithelial cell proliferation / Regulation of PTEN gene transcription / positive regulation of interleukin-8 production / cholesterol homeostasis / regulation of cell growth / TP53 Regulates Metabolic Genes / phosphoprotein binding / cellular response to amino acid stimulus / 加水分解酵素; 酸無水物に作用; GTPに作用・細胞または細胞小器官の運動に関与 / regulation of protein phosphorylation / positive regulation of protein-containing complex assembly / response to virus / MAP2K and MAPK activation / negative regulation of cysteine-type endopeptidase activity involved in apoptotic process / protein localization / negative regulation of ERK1 and ERK2 cascade / mitotic spindle / cilium / positive regulation of protein localization to nucleus / GDP binding 類似検索 - 分子機能
Folliculin / Folliculin, DENN domain / Folliculin, DENN domain, C-terminal superfamily / Folliculin C-terminal domain / Folliculin-interacting protein family / Folliculin-interacting protein, N-terminal domain / Folliculin-interacting protein, middle domain / Folliculin-interacting protein, C-terminal domain / Folliculin-interacting protein N-terminus / Folliculin-interacting protein middle domain ...Folliculin / Folliculin, DENN domain / Folliculin, DENN domain, C-terminal superfamily / Folliculin C-terminal domain / Folliculin-interacting protein family / Folliculin-interacting protein, N-terminal domain / Folliculin-interacting protein, middle domain / Folliculin-interacting protein, C-terminal domain / Folliculin-interacting protein N-terminus / Folliculin-interacting protein middle domain / Folliculin-interacting protein C-terminus / Tripartite DENN domain, FNIP1/2-type / Tripartite DENN FNIP1/2-type domain profile. / Folliculin/SMCR8, longin domain / Folliculin/SMCR8, tripartite DENN domain / Vesicle coat protein involved in Golgi to plasma membrane transport / Tripartite DENN FLCN/SMCR8-type domain profile. / LAMTOR1/MEH1 / Late endosomal/lysosomal adaptor and MAPK and MTOR activator / Late endosomal/lysosomal adaptor and MAPK and MTOR activator / Ragulator complex protein LAMTOR4 / Ragulator complex protein LAMTOR3 / Ragulator complex protein LAMTOR5 / RagA/B / Mitogen-activated protein kinase kinase 1 interacting / Ragulator complex protein LAMTOR5 / Mitogen-activated protein kinase kinase 1 interacting / Gtr1/RagA G protein / RagC/D / Gtr1/RagA G protein conserved region / Ragulator complex protein LAMTOR2-like / Roadblock/LAMTOR2 domain / Roadblock/LC7 domain / Roadblock/LC7 domain / P-loop containing nucleoside triphosphate hydrolase 類似検索 - ドメイン・相同性
Ragulator complex protein LAMTOR5 / Ragulator complex protein LAMTOR5 / Ragulator complex protein LAMTOR4 / Ragulator complex protein LAMTOR1 / Ras-related GTP-binding protein A / Folliculin / Ras-related GTP-binding protein C / Folliculin-interacting protein 2 / Ragulator complex protein LAMTOR3 / Ragulator complex protein LAMTOR2 類似検索 - 構成要素
National Institutes of Health/National Institute of General Medical Sciences (NIH/NIGMS)
2R01GM111730-05
米国
引用
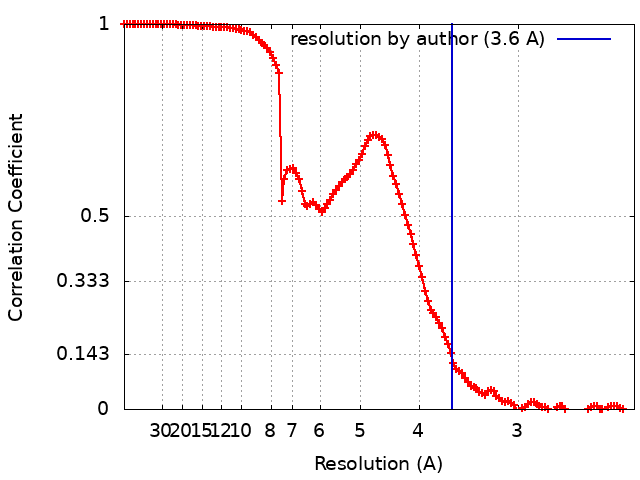
ジャーナル: Science / 年: 2019 タイトル: Structural mechanism of a Rag GTPase activation checkpoint by the lysosomal folliculin complex. 著者: Rosalie E Lawrence / Simon A Fromm / Yangxue Fu / Adam L Yokom / Do Jin Kim / Ashley M Thelen / Lindsey N Young / Chun-Yan Lim / Avi J Samelson / James H Hurley / Roberto Zoncu / 要旨: The tumor suppressor folliculin (FLCN) enables nutrient-dependent activation of the mechanistic target of rapamycin complex 1 (mTORC1) protein kinase via its guanosine triphosphatase (GTPase) ...The tumor suppressor folliculin (FLCN) enables nutrient-dependent activation of the mechanistic target of rapamycin complex 1 (mTORC1) protein kinase via its guanosine triphosphatase (GTPase) activating protein (GAP) activity toward the GTPase RagC. Concomitant with mTORC1 inactivation by starvation, FLCN relocalizes from the cytosol to lysosomes. To determine the lysosomal function of FLCN, we reconstituted the human lysosomal FLCN complex (LFC) containing FLCN, its partner FLCN-interacting protein 2 (FNIP2), and the RagA:RagC GTPases as they exist in the starved state with their lysosomal anchor Ragulator complex and determined its cryo-electron microscopy structure to 3.6 angstroms. The RagC-GAP activity of FLCN was inhibited within the LFC, owing to displacement of a catalytically required arginine in FLCN from the RagC nucleotide. Disassembly of the LFC and release of the RagC-GAP activity of FLCN enabled mTORC1-dependent regulation of the master regulator of lysosomal biogenesis, transcription factor E3, implicating the LFC as a checkpoint in mTORC1 signaling.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 米国, 1件
米国, 1件  引用
引用 構造の表示
構造の表示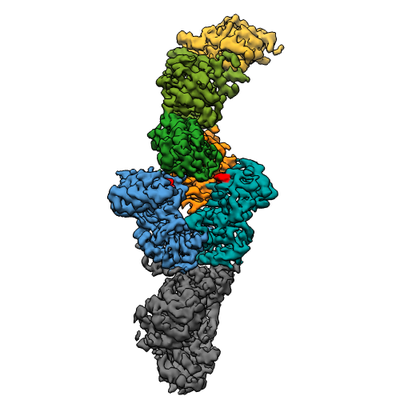
 ダウンロードとリンク
ダウンロードとリンク emd_0554.png
emd_0554.png http://ftp.pdbj.org/pub/emdb/structures/EMD-0554
http://ftp.pdbj.org/pub/emdb/structures/EMD-0554













 Z
Z Y
Y X
X








 試料の構成要素
試料の構成要素
 Spodoptera frugiperda (ツマジロクサヨトウ)
Spodoptera frugiperda (ツマジロクサヨトウ)

 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN