 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-4v7e: Model of the small subunit RNA based on a 5.5 A cryo-EM map of Tr... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 4v7e | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Model of the small subunit RNA based on a 5.5 A cryo-EM map of Triticum aestivum translating 80S ribosome | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | RIBOSOME / eukaryotic ribosome / homology modeling / de novo modeling / ribosomal RNA / rRNA / RNA expansion segments | |||||||||
| Function / homology |  Function and homology information Function and homology informationprotein kinase activator activity / translational elongation / ribonucleoprotein complex binding / maturation of LSU-rRNA / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of SSU-rRNA / small-subunit processome / rRNA processing / ribosomal small subunit biogenesis / ribosomal small subunit assembly ...protein kinase activator activity / translational elongation / ribonucleoprotein complex binding / maturation of LSU-rRNA / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of SSU-rRNA / small-subunit processome / rRNA processing / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / large ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / cytoplasmic translation / negative regulation of translation / ribosome / structural constituent of ribosome / translation / ribonucleoprotein complex / mRNA binding / signal transduction / RNA binding / zinc ion binding / nucleus / cytoplasm / cytosol Similarity search - Function | |||||||||
| Biological species |  | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 5.5 Å | |||||||||
 Authors Authors | Barrio-Garcia, C. / Armache, J.-P. / Jarasch, A. / Anger, A.M. / Villa, E. / Becker, T. / Bhushan, S. / Jossinet, F. / Habeck, M. / Dindar, G. ...Barrio-Garcia, C. / Armache, J.-P. / Jarasch, A. / Anger, A.M. / Villa, E. / Becker, T. / Bhushan, S. / Jossinet, F. / Habeck, M. / Dindar, G. / Franckenberg, S. / Marquez, V. / Mielke, T. / Thomm, M. / Berninghausen, O. / Beatrix, B. / Soeding, J. / Westhof, E. / Wilson, D.N. / Beckmann, R. | |||||||||
 Citation Citation | Journal: Nature / Year: 2014 Title: Structures of the Sec61 complex engaged in nascent peptide translocation or membrane insertion. Authors: Marko Gogala / Thomas Becker / Birgitta Beatrix / Jean-Paul Armache / Clara Barrio-Garcia / Otto Berninghausen / Roland Beckmann /  Abstract: The biogenesis of secretory as well as transmembrane proteins requires the activity of the universally conserved protein-conducting channel (PCC), the Sec61 complex (SecY complex in bacteria). In ...The biogenesis of secretory as well as transmembrane proteins requires the activity of the universally conserved protein-conducting channel (PCC), the Sec61 complex (SecY complex in bacteria). In eukaryotic cells the PCC is located in the membrane of the endoplasmic reticulum where it can bind to translating ribosomes for co-translational protein transport. The Sec complex consists of three subunits (Sec61α, β and γ) and provides an aqueous environment for the translocation of hydrophilic peptides as well as a lateral opening in the Sec61α subunit that has been proposed to act as a gate for the membrane partitioning of hydrophobic domains. A plug helix and a so-called pore ring are believed to seal the PCC against ion flow and are proposed to rearrange for accommodation of translocating peptides. Several crystal and cryo-electron microscopy structures revealed different conformations of closed and partially open Sec61 and SecY complexes. However, in none of these samples has the translocation state been unambiguously defined biochemically. Here we present cryo-electron microscopy structures of ribosome-bound Sec61 complexes engaged in translocation or membrane insertion of nascent peptides. Our data show that a hydrophilic peptide can translocate through the Sec complex with an essentially closed lateral gate and an only slightly rearranged central channel. Membrane insertion of a hydrophobic domain seems to occur with the Sec complex opening the proposed lateral gate while rearranging the plug to maintain an ion permeability barrier. Taken together, we provide a structural model for the basic activities of the Sec61 complex as a protein-conducting channel. #1: Journal: Proc Natl Acad Sci U S A / Year: 2010Title: Cryo-EM structure and rRNA model of a translating eukaryotic 80S ribosome at 5.5-A resolution. Authors: Jean-Paul Armache / Alexander Jarasch / Andreas M Anger / Elizabeth Villa / Thomas Becker / Shashi Bhushan / Fabrice Jossinet / Michael Habeck / Gülcin Dindar / Sibylle Franckenberg / Viter ...Authors: Jean-Paul Armache / Alexander Jarasch / Andreas M Anger / Elizabeth Villa / Thomas Becker / Shashi Bhushan / Fabrice Jossinet / Michael Habeck / Gülcin Dindar / Sibylle Franckenberg / Viter Marquez / Thorsten Mielke / Michael Thomm / Otto Berninghausen / Birgitta Beatrix / Johannes Söding / Eric Westhof / Daniel N Wilson / Roland Beckmann / Abstract: Protein biosynthesis, the translation of the genetic code into polypeptides, occurs on ribonucleoprotein particles called ribosomes. Although X-ray structures of bacterial ribosomes are available, ...Protein biosynthesis, the translation of the genetic code into polypeptides, occurs on ribonucleoprotein particles called ribosomes. Although X-ray structures of bacterial ribosomes are available, high-resolution structures of eukaryotic 80S ribosomes are lacking. Using cryoelectron microscopy and single-particle reconstruction, we have determined the structure of a translating plant (Triticum aestivum) 80S ribosome at 5.5-Å resolution. This map, together with a 6.1-Å map of a Saccharomyces cerevisiae 80S ribosome, has enabled us to model ∼98% of the rRNA. Accurate assignment of the rRNA expansion segments (ES) and variable regions has revealed unique ES-ES and r-protein-ES interactions, providing insight into the structure and evolution of the eukaryotic ribosome. #2: Journal: Proc Natl Acad Sci U S A / Year: 2010Title: Localization of eukaryote-specific ribosomal proteins in a 5.5-Å cryo-EM map of the 80S eukaryotic ribosome. Authors: Jean-Paul Armache / Alexander Jarasch / Andreas M Anger / Elizabeth Villa / Thomas Becker / Shashi Bhushan / Fabrice Jossinet / Michael Habeck / Gülcin Dindar / Sibylle Franckenberg / Viter ...Authors: Jean-Paul Armache / Alexander Jarasch / Andreas M Anger / Elizabeth Villa / Thomas Becker / Shashi Bhushan / Fabrice Jossinet / Michael Habeck / Gülcin Dindar / Sibylle Franckenberg / Viter Marquez / Thorsten Mielke / Michael Thomm / Otto Berninghausen / Birgitta Beatrix / Johannes Söding / Eric Westhof / Daniel N Wilson / Roland Beckmann / Abstract: Protein synthesis in all living organisms occurs on ribonucleoprotein particles, called ribosomes. Despite the universality of this process, eukaryotic ribosomes are significantly larger in size than ...Protein synthesis in all living organisms occurs on ribonucleoprotein particles, called ribosomes. Despite the universality of this process, eukaryotic ribosomes are significantly larger in size than their bacterial counterparts due in part to the presence of 80 r proteins rather than 54 in bacteria. Using cryoelectron microscopy reconstructions of a translating plant (Triticum aestivum) 80S ribosome at 5.5-Å resolution, together with a 6.1-Å map of a translating Saccharomyces cerevisiae 80S ribosome, we have localized and modeled 74/80 (92.5%) of the ribosomal proteins, encompassing 12 archaeal/eukaryote-specific small subunit proteins as well as the complete complement of the ribosomal proteins of the eukaryotic large subunit. Near-complete atomic models of the 80S ribosome provide insights into the structure, function, and evolution of the eukaryotic translational apparatus. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 4v7e.cif.gz | 4.7 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb4v7e.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 4v7e.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 4v7e_validation.pdf.gz | 1.8 MB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 4v7e_full_validation.pdf.gz | 3.4 MB | Display | |
| Data in XML | 4v7e_validation.xml.gz | 474.5 KB | Display | |
| Data in CIF | 4v7e_validation.cif.gz | 769.6 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/v7/4v7eftp://data.pdbj.org/pub/pdb/validation_reports/v7/4v7e | HTTPS FTP |
-Related structure data
| Related structure data |  1780M  2510C  2511C  2512C  4cg5C  4cg6C  4cg7C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |





-Links
 PDBj
PDBj






























- Assembly
Assembly
| Deposited unit | 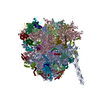
|
|---|---|
| 1 |
|
-Components
-RNA chain , 6 types, 6 molecules AdAeAfAaAcAb
| #1: RNA chain | Mass: 583613.562 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #2: RNA chain | Mass: 24135.262 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #3: RNA chain | Mass: 3498.178 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Details: messenger RNA / Source: (natural) |
| #84: RNA chain | Mass: 1097289.250 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #85: RNA chain | Mass: 51535.613 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #86: RNA chain | Mass: 38716.957 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
+40S ribosomal protein ... , 32 types, 32 molecules BYBIBKBMBfBXBDBEBFBQBUBOBSBNBLBTBPBZBcBWBdBbBeBABRBBBVBaBJBCBGBH
-Protein , 3 types, 4 molecules BgCsCtCq
| #10: Protein | Mass: 41794.809 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||
|---|---|---|---|
| #67: Protein | Mass: 11596.995 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #70: Protein | | Mass: 34418.574 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
+60S ribosomal protein ... , 45 types, 46 molecules CGCTCZCzCACJCHCVCNCaCQCDCRCPCXCWCYCrCcCdCeCjClCoCMCSCUCiCKCu...
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 4.2 MDa / Experimental value: YES | ||||||||||||
| Buffer solution | Name: 20 mM HEPES/KOH, pH 7.5, 100 mM KOAc, 10 mM Mg(OAc)2, 0.01 mg/mL cycloheximide, 1 mM DTT, 0.01% Nikkol pH: 7.5 Details: 20 mM HEPES/KOH, pH 7.5, 100 mM KOAc, 10 mM Mg(OAc)2, 0.01 mg/mL cycloheximide, 1 mM DTT, 0.01% Nikkol | ||||||||||||
| Specimen | Conc.: 0.02 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||
| Specimen support | Details: Quantifoil Grid with 2 nm carbon on top | ||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK I / Cryogen name: ETHANE / Humidity: 100 % Details: Blot for 10 seconds (using 2 layers of filter paper) before plunging into liquid ethane (FEI VITROBOT). Method: Blot for 10 seconds before plunging, use 2 layers of filter paper |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Tecnai F30 / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TECNAI F30 |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 39000 X / Calibrated magnification: 38900 X / Nominal defocus max: 4500 nm / Nominal defocus min: 1000 nm / Cs: 2.26 mm |
| Specimen holder | Specimen holder model: OTHER / Specimen holder type: FEI Polara Cartridge System |
| Image recording | Electron dose: 25 e/Å2 / Film or detector model: KODAK SO-163 FILM Details: Scanned at 5334 dpi on a Heidelberg Primescan Drum Scanner |
| Image scans | Num. digital images: 1374 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Relative weight: 1 |
- Processing
Processing
| EM software | Name: SPIDER / Category: 3D reconstruction | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Details: Wiener Filter on 3D volumes (SPIDER) | ||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||
| 3D reconstruction | Method: Projection Matching / Resolution: 5.5 Å / Resolution method: FSC 0.5 CUT-OFF / Num. of particles: 2108230 / Nominal pixel size: 1.24 Å / Actual pixel size: 1.24 Å / Symmetry type: POINT | ||||||||||||
| Refinement step | Cycle: LAST
|
