 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 2wpd | ||||||
|---|---|---|---|---|---|---|---|
| Title | The Mg.ADP inhibited state of the yeast F1c10 ATP synthase | ||||||
 Components Components | (ATP SYNTHASE SUBUNIT ... ) x 6 ) x 6 | ||||||
 Keywords Keywords | HYDROLASE / ATP PHOSPHORYLASE (H+ TRANSPORTING) / ATP-BINDING / CENTRAL STALK / ATP SYNTHESIS / PHOSPHOPROTEIN / MEMBRANE PROTEIN / LIPID-BINDING / ION TRANSPORT / NUCLEOTIDE-BINDING / HYDROGEN ION TRANSPORT | ||||||
| Function / homology |  Function and homology information Function and homology informationmitochondrial proton-transporting ATP synthase, central stalk / mitochondrial proton-transporting ATP synthase, catalytic core / mitochondrial proton-transporting ATP synthase complex, coupling factor F(o) / mitochondrial proton-transporting ATP synthase complex / mitochondrial proton-transporting ATP synthase complex, catalytic sector F(1) / mitochondrial nucleoid / proton motive force-driven mitochondrial ATP synthesis / proton motive force-driven ATP synthesis / proton transmembrane transporter activity / proton-transporting ATP synthase complex, catalytic core F(1) ...mitochondrial proton-transporting ATP synthase, central stalk / mitochondrial proton-transporting ATP synthase, catalytic core / mitochondrial proton-transporting ATP synthase complex, coupling factor F(o) / mitochondrial proton-transporting ATP synthase complex / mitochondrial proton-transporting ATP synthase complex, catalytic sector F(1) / mitochondrial nucleoid / proton motive force-driven mitochondrial ATP synthesis / proton motive force-driven ATP synthesis / proton transmembrane transporter activity / proton-transporting ATP synthase complex, catalytic core F(1) / H+-transporting two-sector ATPase / proton-transporting ATPase activity, rotational mechanism / proton-transporting ATP synthase activity, rotational mechanism / ADP binding / mitochondrial intermembrane space / mitochondrial inner membrane / lipid binding / ATP hydrolysis activity / mitochondrion / ATP binding / identical protein binding / cytosolSimilarity search - Function | ||||||
| Biological species |  SACCHAROMYCES CEREVISIAE (brewer's yeast) SACCHAROMYCES CEREVISIAE (brewer's yeast) | ||||||
| Method | X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 3.432 Å | ||||||
 Authors Authors | Dautant, A. / Velours, J. / Giraud, M.-F. | ||||||
 Citation Citation | Journal: J.Biol.Chem. / Year: 2010 Title: Crystal Structure of the Mg.Adp-Inhibited State of the Yeast F1C10-ATP Synthase. Authors: Dautant, A. / Velours, J. / Giraud, M. #1: Journal: Science / Year: 1999Title: Molecular Architecture of the Rotary Motor in ATP Synthase. Authors: Stock, D. / Leslie, A.G.W. / Walker, J.E. #2: Journal: Embo J. / Year: 2006Title: Novel Features of the Rotary Catalytic Mechanism Revealed in the Structure of Yeast F1 ATPase. Authors: Kabaleeswaran, V. / Puri, N. / Walker, J.E. / Leslie, A.G.W. / Mueller, D.M. #3: Journal: J.Biol.Chem. / Year: 2009Title: Asymmetric Structure of the Yeast F1 ATPase in the Absence of Bound Nucleotides. Authors: Kabaleeswaran, V. / Shen, H. / Symersky, J. / Walker, J.E. / Leslie, A.G.W. / Mueller, D.M. #4: Journal: Science / Year: 2005Title: Structure of the Rotor Ring of F-Type Na+-ATPase from Ilyobacter Tartaricus. Authors: Meier, T. / Polzer, P. / Diederichs, K. / Welte, W. / Dimroth, P. #5: Journal: J.Bioenerg.Biomembr. / Year: 2009 Title: Hydrogenated and Fluorinated Surfactants Derived from Tris(Hydroxymethyl)-Acrylamidomethane Allow the Purification of a Highly Active Yeast F1-F0 ATP-Synthase with an Enhanced Stability. Authors: Talbot, J.-C. / Dautant, A. / Polidori, A. / Pucci, B. / Cohen-Bouhacina, T. / Maali, A. / Salin, B. / Brethes, D. / Velours, J. / Giraud, M.-F. | ||||||
| History |
| ||||||
| Remark 650 | HELIX DETERMINATION METHOD: AUTHOR PROVIDED. |
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 2wpd.cif.gz | 656.1 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb2wpd.ent.gz | 524.8 KB | Display | PDB format |
| PDBx/mmJSON format | 2wpd.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/wp/2wpdftp://data.pdbj.org/pub/pdb/validation_reports/wp/2wpd | HTTPS FTP |
|---|
-Related structure data







-Links
 PDBj
PDBj
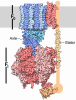

- Assembly
Assembly
| Deposited unit | 
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Unit cell |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Noncrystallographic symmetry (NCS) | NCS domain:
NCS domain segments:
|
-Components
-ATP SYNTHASE SUBUNIT ... , 6 types, 19 molecules ABCDEFGHIJKLMNOPQRS
| #1: Protein | / ATP SYNTHASE ALPHA CHAIN\ / MITOCHONDRIAL Mass: 55007.402 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Source: (natural) SACCHAROMYCES CEREVISIAE (brewer's yeast) / Strain: D273-10B/A / References: UniProt: P07251#2: Protein | / ATP SYNTHASE BETA CHAIN\ / MITOCHONDRIALMass: 51181.082 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Source: (natural) SACCHAROMYCES CEREVISIAE (brewer's yeast) / Strain: D273-10B/AReferences: UniProt: P00830, H+-transporting two-sector ATPase#3: Protein | | / ATP SYNTHASE GAMMA CHAIN\ / MITOCHONDRIAL / F-ATPASE GAMMA SUBUNITMass: 30657.160 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) SACCHAROMYCES CEREVISIAE (brewer's yeast) / Strain: D273-10B/A / References: UniProt: P38077#4: Protein | | / ATP SYNTHASE DELTA CHAIN\ / MITOCHONDRIAL / F-ATPASE DELTA SUBUNITMass: 14565.385 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) SACCHAROMYCES CEREVISIAE (brewer's yeast) / Strain: D273-10B/A / References: UniProt: Q12165#5: Protein | | / ATP SYNTHASE EPSILON CHAIN\ / MITOCHONDRIALMass: 6618.359 Da / Num. of mol.: 1 / Fragment: RESIDUES 2-62 / Source method: isolated from a natural source / Source: (natural) SACCHAROMYCES CEREVISIAE (brewer's yeast) / Strain: D273-10B/A / References: UniProt: P21306#6: Protein | / ATP SYNTHASE 9 CHAIN\ / MITOCHONDRIAL / LIPID-BINDING PROTEIN / OLIGOMYCIN RESISTANCE PROTEIN 1Mass: 7762.375 Da / Num. of mol.: 10 / Source method: isolated from a natural source / Source: (natural) SACCHAROMYCES CEREVISIAE (brewer's yeast) / Strain: D273-10B/AReferences: UniProt: P61829, H+-transporting two-sector ATPase |
|---|
-Non-polymers , 3 types, 10 molecules 


| #7: Chemical | Adenosine triphosphate Mass: 507.181 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM#8: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 5 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 5 / Source method: obtained synthetically / Formula: Mg#9: Chemical | Adenosine diphosphate Mass: 427.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 2 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 3.72 Å3/Da / Density % sol: 66.67 % / Description: NONE |
|---|---|
| Crystal grow | pH: 7.5 Details: 0.1 M HEPES/HCL PH 7.5, 12% PEG MME 5000, 100 MM SODIUM CHLORIDE MIXED 1:1 WITH PROTEIN SOLUTION CONTAINING 0.64 MM DDM, 25 MM TRIS/HCL PH 8.0, 100 MM SODIUM CHLORIDE, 25 MM TREHALOSE, 0.5 ...Details: 0.1 M HEPES/HCL PH 7.5, 12% PEG MME 5000, 100 MM SODIUM CHLORIDE MIXED 1:1 WITH PROTEIN SOLUTION CONTAINING 0.64 MM DDM, 25 MM TRIS/HCL PH 8.0, 100 MM SODIUM CHLORIDE, 25 MM TREHALOSE, 0.5 MM EDTA, 0.02% SODIUM AZIDE, 2 MM MAGNESIUM CHLORIDE, 0.66 MM ADP, 0.1 MM DCCD, 2.5 MM DTT, 0.5 MM PMSF |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: ESRF  / Beamline: ID29 / Wavelength: 1 / Beamline: ID29 / Wavelength: 1 |
| Detector | Type: MARRESEARCH / Detector: CCD / Date: Feb 4, 2005 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1 Å / Relative weight: 1 |
| Reflection | Resolution: 3.43→39.8 Å / Num. obs: 84424 / % possible obs: 88.1 % / Observed criterion σ(I): 0 / Redundancy: 2.8 % / Biso Wilson estimate: 85 Å2 / Rmerge(I) obs: 0.19 / Net I/σ(I): 4.3 |
| Reflection shell | Resolution: 3.43→3.62 Å / Redundancy: 2.3 % / Rmerge(I) obs: 0.72 / Mean I/σ(I) obs: 2.3 / % possible all: 64.4 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: PDB ENTRIES 2HLD, 1YCE Resolution: 3.432→39.775 Å / SU ML: 0.67 / σ(F): 1.34 / Phase error: 32.75 / Stereochemistry target values: ML
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.8 Å / VDW probe radii: 0.8 Å / Solvent model: FLAT BULK SOLVENT MODEL / Bsol: 28.559 Å2 / ksol: 0.281 e/Å3 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 108 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 3.432→39.775 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints NCS |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
