 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6pwb | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
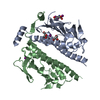
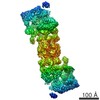
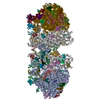
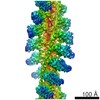
| タイトル | Rigid body fitting of flagellin FlaB, and flagellar coiling proteins, FcpA and FcpB, into a 10 Angstrom structure of the asymmetric flagellar filament purified from Leptospira biflexa Patoc WT cells resolved via subtomogram averaging | |||||||||||||||||||||||||||||||||
 要素 要素 |
| |||||||||||||||||||||||||||||||||
 キーワード キーワード |  STRUCTURAL PROTEIN (タンパク質) / bacterial flagella (鞭毛) / FcpA / FcpB / FlaA / FlaB / Leptospira (レプトスピラ属) STRUCTURAL PROTEIN (タンパク質) / bacterial flagella (鞭毛) / FcpA / FcpB / FlaA / FlaB / Leptospira (レプトスピラ属) | |||||||||||||||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報periplasmic flagellum / bacterial-type flagellum filament / bacterial-type flagellum-dependent cell motility / structural molecule activity 類似検索 - 分子機能 | |||||||||||||||||||||||||||||||||
| 生物種 |  Leptospira biflexa serovar Patoc (バクテリア) Leptospira biflexa serovar Patoc (バクテリア) | |||||||||||||||||||||||||||||||||
| 手法 | 電子顕微鏡法 / サブトモグラム平均法 / クライオ電子顕微鏡法 / 解像度: 9.83 Å | |||||||||||||||||||||||||||||||||
 データ登録者 データ登録者 | Gibson, K.H. / Sindelar, C.V. / Trajtenberg, F. / Buschiazzo, A. / San Martin, F. / Mechaly, A. | |||||||||||||||||||||||||||||||||
| 資金援助 |  米国, ウルグアイ, 10件 米国, ウルグアイ, 10件
| |||||||||||||||||||||||||||||||||
 引用 引用 | ジャーナル: Elife / 年: 2020 タイトル: An asymmetric sheath controls flagellar supercoiling and motility in the leptospira spirochete. 著者: Kimberley H Gibson / Felipe Trajtenberg / Elsio A Wunder / Megan R Brady / Fabiana San Martin / Ariel Mechaly / Zhiguo Shang / Jun Liu / Mathieu Picardeau / Albert Ko / Alejandro Buschiazzo / ...著者: Kimberley H Gibson / Felipe Trajtenberg / Elsio A Wunder / Megan R Brady / Fabiana San Martin / Ariel Mechaly / Zhiguo Shang / Jun Liu / Mathieu Picardeau / Albert Ko / Alejandro Buschiazzo / Charles Vaughn Sindelar /   要旨: Spirochete bacteria, including important pathogens, exhibit a distinctive means of swimming via undulations of the entire cell. Motility is powered by the rotation of supercoiled 'endoflagella' that ...Spirochete bacteria, including important pathogens, exhibit a distinctive means of swimming via undulations of the entire cell. Motility is powered by the rotation of supercoiled 'endoflagella' that wrap around the cell body, confined within the periplasmic space. To investigate the structural basis of flagellar supercoiling, which is critical for motility, we determined the structure of native flagellar filaments from the spirochete by integrating high-resolution cryo-electron tomography and X-ray crystallography. We show that these filaments are coated by a highly asymmetric, multi-component sheath layer, contrasting with flagellin-only homopolymers previously observed in exoflagellated bacteria. Distinct sheath proteins localize to the filament inner and outer curvatures to define the supercoiling geometry, explaining a key functional attribute of this spirochete flagellum. #1: ジャーナル: Acta Crystallogr F Struct Biol Commun / 年: 2017 タイトル: Crystallization of FcpA from Leptospira, a novel flagellar protein that is essential for pathogenesis. 著者: Fabiana San Martin / Ariel E Mechaly / Nicole Larrieux / Elsio A Wunder / Albert I Ko / Mathieu Picardeau / Felipe Trajtenberg / Alejandro Buschiazzo / 要旨: The protein FcpA is a unique component of the flagellar filament of spirochete bacteria belonging to the genus Leptospira. Although it plays an essential role in translational motility and ...The protein FcpA is a unique component of the flagellar filament of spirochete bacteria belonging to the genus Leptospira. Although it plays an essential role in translational motility and pathogenicity, no structures of FcpA homologues are currently available in the PDB. Its three-dimensional structure will unveil the novel motility mechanisms that render pathogenic Leptospira particularly efficient at invading and disseminating within their hosts, causing leptospirosis in humans and animals. FcpA from L. interrogans was purified and crystallized, but despite laborious attempts no useful X ray diffraction data could be obtained. This challenge was solved by expressing a close orthologue from the related saprophytic species L. biflexa. Three different crystal forms were obtained: a primitive and a centred monoclinic form, as well as a hexagonal variant. All forms diffracted X-rays to suitable resolutions for crystallographic analyses, with the hexagonal type typically reaching the highest limits of 2.0 Å and better. A variation of the quick-soaking procedure resulted in an iodide derivative that was instrumental for single-wavelength anomalous diffraction methods. #2: ジャーナル: Mol Microbiol / 年: 2016 タイトル: A novel flagellar sheath protein, FcpA, determines filament coiling, translational motility and virulence for the Leptospira spirochete. 著者: Elsio A Wunder / Cláudio P Figueira / Nadia Benaroudj / Bo Hu / Brian A Tong / Felipe Trajtenberg / Jun Liu / Mitermayer G Reis / Nyles W Charon / Alejandro Buschiazzo / Mathieu Picardeau / Albert I Ko / 要旨: Leptospira are unique among bacteria based on their helical cell morphology with hook-shaped ends and the presence of periplasmic flagella (PF) with pronounced spontaneous supercoiling. The factors ...Leptospira are unique among bacteria based on their helical cell morphology with hook-shaped ends and the presence of periplasmic flagella (PF) with pronounced spontaneous supercoiling. The factors that provoke such supercoiling, as well as the role that PF coiling plays in generating the characteristic hook-end cell morphology and motility, have not been elucidated. We have now identified an abundant protein from the pathogen L. interrogans, exposed on the PF surface, and named it Flagellar-coiling protein A (FcpA). The gene encoding FcpA is highly conserved among Leptospira and was not found in other bacteria. fcpA(-) mutants, obtained from clinical isolates or by allelic exchange, had relatively straight, smaller-diameter PF, and were not able to produce translational motility. These mutants lost their ability to cause disease in the standard hamster model of leptospirosis. Complementation of fcpA restored the wild-type morphology, motility and virulence phenotypes. In summary, we identified a novel Leptospira 36-kDa protein, the main component of the spirochete's PF sheath, and a key determinant of the flagella's coiled structure. FcpA is essential for bacterial translational motility and to enable the spirochete to penetrate the host, traverse tissue barriers, disseminate to cause systemic infection and reach target organs. #3: ジャーナル: Front Cell Infect Microbiol / 年: 2018 タイトル: FcpB Is a Surface Filament Protein of the Endoflagellum Required for the Motility of the Spirochete . 著者: Elsio A Wunder / Leyla Slamti / David N Suwondo / Kimberley H Gibson / Zhiguo Shang / Charles V Sindelar / Felipe Trajtenberg / Alejandro Buschiazzo / Albert I Ko / Mathieu Picardeau / 要旨: The spirochete endoflagellum is a unique motility apparatus among bacteria. Despite its critical importance for pathogenesis, the full composition of the flagellum remains to be determined. We have ...The spirochete endoflagellum is a unique motility apparatus among bacteria. Despite its critical importance for pathogenesis, the full composition of the flagellum remains to be determined. We have recently reported that FcpA is a novel flagellar protein and a major component of the sheath of the filament of the spirochete . By screening a library of random transposon mutants in the spirochete , we found a motility-deficient mutant harboring a disruption in a hypothetical gene of unknown function. Here, we show that this gene encodes a surface component of the endoflagellar filament and is required for typical hook- and spiral-shaped ends of the cell body, coiled structure of the endoflagella, and high velocity phenotype. We therefore named the gene for flagellar-coiling protein B. is conserved in all members of the genus, but not present in other organisms including other spirochetes. Complementation of the mutant restored the wild-type morphology and motility phenotypes. Immunoblotting with anti-FcpA and anti-FcpB antisera and cryo-electron microscopy of the filament indicated that FcpB assembled onto the surface of the sheath of the filament and mostly located on the outer (convex) side of the coiled filament. We provide evidence that FcpB, together with FcpA, are -specific novel components of the sheath of the filament, key determinants of the coiled and asymmetric structure of the endoflagella and are essential for high velocity. Defining the components of the endoflagella and their functions in these atypical bacteria should greatly enhance our understanding of the mechanisms by which these bacteria produce motility. | |||||||||||||||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6pwb.cif.gz | 4.5 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6pwb.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 6pwb.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/pw/6pwbftp://data.pdbj.org/pub/pdb/validation_reports/pw/6pwb | HTTPS FTP |
|---|
-関連構造データ










-リンク
 PDBj
PDBj- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 29467.033 Da / 分子数: 84 / 由来タイプ: 天然 由来: (天然) Leptospira biflexa serovar Patoc (strain Patoc 1 / ATCC 23582 / Paris) (バクテリア)Cell: bacteria / 株: Patoc 1 / ATCC 23582 / Paris / 参照: UniProt: B0SSZ5 #2: タンパク質 | 分子量: 28931.002 Da / 分子数: 47 / 由来タイプ: 天然 由来: (天然) Leptospira biflexa serovar Patoc (strain Patoc 1 / ATCC 23582 / Paris) (バクテリア)Cell: bacteria / 株: Patoc 1 / ATCC 23582 / Paris / 参照: UniProt: B0STJ8 #3: タンパク質 | 分子量: 25539.666 Da / 分子数: 32 / 由来タイプ: 天然 由来: (天然) Leptospira biflexa serovar Patoc (strain Patoc 1 / ATCC 23582 / Paris) (バクテリア)Cell: bacteria / 株: Patoc 1 / ATCC 23582 / Paris / 参照: UniProt: B0SR03 |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: FILAMENT / 3次元再構成法: サブトモグラム平均法 |
- 試料調製
試料調製
| 構成要素 | 名称: Flagellar filament purified from the periplasm of the Spirochete bacterium, Leptospira biflexa タイプ: ORGANELLE OR CELLULAR COMPONENT 詳細: Filaments were purified from Leptospira biflexa wild type cells. Entity ID: all / 由来: NATURAL |
|---|---|
| 分子量 | 実験値: NO |
| 由来(天然) | 生物種: Leptospira biflexa serovar Patoc strain 'Patoc 1 (Paris)' (バクテリア) 細胞内の位置: periplasm / Organelle : flagellar filament |
| 緩衝液 | pH: 6.8 / 詳細: Tris-HCl or ddH20, pH6.8 with <1% sodium azide as a preservative. FIDUCIAL MARKERS: Prior to vitrification, 1 uL of 6x concentrated Gold Tracer beads (10 nm colloidal gold) were mixed with 2 uL of purified flagella. To each grid, 3 uL of this mixture were applied. |
| 試料 | 濃度: 0.5 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES 詳細: Leptospira biflexa serovar Patoc strain Patoc I wild type, fcpA, and fcpB mutant cells cultured in Ellinghausen-McCullough-Johnson-Harris liquid medium until they reached logarithmic phase at ...詳細: Leptospira biflexa serovar Patoc strain Patoc I wild type, fcpA, and fcpB mutant cells cultured in Ellinghausen-McCullough-Johnson-Harris liquid medium until they reached logarithmic phase at 30C. The cells were pelleted and the periplasmic flagellar filaments were purified as described in Wunder et al., 2016; Wunder et al., 2018. |
| 試料支持 | 詳細: Model 950 Solarus Advanced Plasma System manufactured by GATAN. グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R1.2/1.3 |
| 急速凍結 | 装置: FEI VITROBOT MARK III / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 291 K 詳細: 2 minute incubation time; blot time of 6-7.5 seconds; and blot offset of -2 mm |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Tecnai Polara / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI POLARA 300 |
| 電子銃 | 電子線源: FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELDBright-field microscopy / 倍率(公称値): 15000 X / 倍率(補正後): 19230 X / 最大 デフォーカス(公称値): 5000 nm / 最小 デフォーカス(公称値): 3000 nm / Cs: 2 mm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN / 試料ホルダーモデル: SIDE ENTRY, EUCENTRIC |
| 撮影 | 平均露光時間: 1.2 sec. / 電子線照射量: 1.7 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 1 詳細: Tilt series acquisition. A total of ~35 tilt angles per tilt stack were acquired. The total dose was ~ 60 e/Angstrom2. |
| 画像スキャン | サンプリングサイズ: 5 µm / 横: 3710 / 縦: 3838 / 動画フレーム数/画像: 12 / 利用したフレーム数/画像: 1-12 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 画像処理 | 詳細: Motion correction was performed on movie stacks acquired at each angle in each tilt series using IMOD alignframes. Tilt series were aligned in IMOD eTomo usinf 10 nm fiducial gold markers. ...詳細: Motion correction was performed on movie stacks acquired at each angle in each tilt series using IMOD alignframes. Tilt series were aligned in IMOD eTomo usinf 10 nm fiducial gold markers. CTF estimation with phase flipping was carried out, along with gold bead subtraction in IMOD eTomo. Tilt series were reconstructed using Weighted Back Projection in Tomo3D. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | 詳細: CTF estimation was initially done in IMOD. / タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 9.83 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 10581 / アルゴリズム: BACK PROJECTION 詳細: emClarity: The two half-dataset volumes were combined with a B-170 factor of 0, and the Gold standard FSC at 0.143 was calculated to be 9.83 A with anisotropic resolutions ranging from 8.89-16.17 Angstrom. 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EM volume selection | 手法: Manual selection 詳細: Using IMOD 3dmod, filament trajectories were traced by selecting particle points along a single filament. Each continuous filament was a single contour (3dmod). Using the addModPts program in ...詳細: Using IMOD 3dmod, filament trajectories were traced by selecting particle points along a single filament. Each continuous filament was a single contour (3dmod). Using the addModPts program in PEET, particle gaps along each filament trajectory contour were filled in with additional points according to a repeat spacing of 52 angstroms. The x,y,z coordinates were then imported into RELION using a modified version of the RELION preprocessing.py python script described in Bharat et. al. (2015). The script was modified to ensure that particles in one filament would be sorted into the same ODD or EVEN grouping to prevent over-estimation of FSC due to inclusion of the 2 or more overlapping particles in a filament in both half-datasets. Num. of tomograms: 62 / Num. of volumes extracted: 15000 / Reference model: de novo | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: RECIPROCAL / Target criteria: Correlation coefficient 詳細: Eliminate minor clashes between FlaB1, FcpA, and FcpB | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
|
