ornithine decarboxylase inhibitor activity / transcription antitermination factor activity, RNA binding / misfolded RNA binding / Group I intron splicing / RNA folding / four-way junction DNA binding / negative regulation of translational initiation / regulation of mRNA stability / mRNA regulatory element binding translation repressor activity / positive regulation of RNA splicing ...ornithine decarboxylase inhibitor activity / transcription antitermination factor activity, RNA binding / misfolded RNA binding / Group I intron splicing / RNA folding / four-way junction DNA binding / negative regulation of translational initiation / regulation of mRNA stability / mRNA regulatory element binding translation repressor activity / positive regulation of RNA splicing / transcription elongation factor complex / regulation of DNA-templated transcription elongation / DNA endonuclease activity / transcription antitermination / DNA-templated transcription termination / maintenance of translational fidelity / mRNA 5'-UTR binding / regulation of translation / ribosome biogenesis / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / cytosolic small ribosomal subunit / small ribosomal subunit rRNA binding / cytoplasmic translation / tRNA binding / rRNA binding / ribosome / structural constituent of ribosome / translation / response to antibiotic / mRNA binding / RNA binding / zinc ion binding / cytosol / cytoplasm 類似検索 - 分子機能
Ribosomal protein S14, bacterial/plastid / Ribosomal protein S16, conserved site / Ribosomal protein S16 signature. / Ribosomal protein S19, bacterial-type / Ribosomal protein S3, bacterial-type / Ribosomal protein S13, bacterial-type / Ribosomal protein S6, conserved site / Ribosomal protein S6 signature. / Ribosomal protein S9, bacterial/plastid / Ribosomal protein S11, bacterial-type ...Ribosomal protein S14, bacterial/plastid / Ribosomal protein S16, conserved site / Ribosomal protein S16 signature. / Ribosomal protein S19, bacterial-type / Ribosomal protein S3, bacterial-type / Ribosomal protein S13, bacterial-type / Ribosomal protein S6, conserved site / Ribosomal protein S6 signature. / Ribosomal protein S9, bacterial/plastid / Ribosomal protein S11, bacterial-type / Ribosomal protein S20 / Ribosomal protein S20 superfamily / Ribosomal protein S20 / Ribosomal protein S4, bacterial-type / 30S ribosomal protein S17 / Ribosomal protein S5, bacterial-type / Ribosomal protein S18, conserved site / Ribosomal protein S18 signature. / Ribosomal protein S6, plastid/chloroplast / Ribosomal protein S16 / Ribosomal protein S16 domain superfamily / Ribosomal protein S16 / Ribosomal protein S15, bacterial-type / Ribosomal protein S12, bacterial-type / Ribosomal protein S18 / Ribosomal protein S18 / Ribosomal protein S18 superfamily / K Homology domain / K homology RNA-binding domain / Ribosomal protein S6 / Ribosomal protein S6 / Ribosomal protein S6 superfamily / Translation elongation factor EF1B/ribosomal protein S6 / Ribosomal protein S3, conserved site / Ribosomal protein S3 signature. / Ribosomal protein S10, conserved site / Ribosomal protein S10 signature. / Ribosomal protein S14, conserved site / Ribosomal protein S14 signature. / KH domain / Type-2 KH domain profile. / K Homology domain, type 2 / Ribosomal protein S3, C-terminal / Ribosomal protein S3, C-terminal domain / Ribosomal protein S3, C-terminal domain superfamily / Ribosomal protein S15/S19, conserved site / Ribosomal protein S19 signature. / Ribosomal protein S10 / Ribosomal protein S19/S15 / Ribosomal protein S19/S15, superfamily / Ribosomal protein S19 / : / Ribosomal protein S5, N-terminal, conserved site / Ribosomal protein S5 signature. / Ribosomal protein S17, conserved site / Ribosomal protein S17 signature. / K homology domain superfamily, prokaryotic type / Ribosomal protein S5 / Ribosomal protein S13, conserved site / Ribosomal protein S13 signature. / S5 double stranded RNA-binding domain profile. / Ribosomal protein S5, N-terminal / Ribosomal protein S13 / 30s ribosomal protein S13, C-terminal / Ribosomal protein S13/S18 / Ribosomal protein S5, C-terminal / Ribosomal protein S13 family profile. / Ribosomal protein S5, N-terminal domain / Ribosomal protein S5, C-terminal domain / Ribosomal protein S8 signature. / Ribosomal protein S4/S9 N-terminal domain / Ribosomal protein S4, conserved site / Ribosomal protein S15 signature. / Ribosomal protein S4 signature. / Ribosomal protein S4/S9 N-terminal domain / Ribosomal protein S4/S9, N-terminal / Ribosomal protein S14 / Ribosomal protein S14p/S29e / Ribosomal protein S4/S9 / K homology domain-like, alpha/beta / Ribosomal protein S8 / Ribosomal protein S8 superfamily / Ribosomal protein S8 / S4 RNA-binding domain profile. / Ribosomal protein S13-like, H2TH / Ribosomal S11, conserved site / Ribosomal protein S11 signature. / S4 RNA-binding domain / S4 domain / RNA-binding S4 domain / Ribosomal protein S10p/S20e / Ribosomal protein S9, conserved site / Ribosomal protein S9 signature. / Ribosomal protein S11 / Ribosomal protein S10 domain / Ribosomal protein S10 domain superfamily / Ribosomal protein S10p/S20e / RNA-binding S4 domain superfamily / Ribosomal protein S11 / Ribosomal protein S12 signature. 類似検索 - ドメイン・相同性
30S ribosomal protein S15 / Small ribosomal subunit protein bS6 / Small ribosomal subunit protein uS10 / Small ribosomal subunit protein uS11 / Small ribosomal subunit protein uS12 / Small ribosomal subunit protein uS13 / Small ribosomal subunit protein bS16 / Small ribosomal subunit protein bS18 / Small ribosomal subunit protein uS19 / Small ribosomal subunit protein bS20 ...30S ribosomal protein S15 / Small ribosomal subunit protein bS6 / Small ribosomal subunit protein uS10 / Small ribosomal subunit protein uS11 / Small ribosomal subunit protein uS12 / Small ribosomal subunit protein uS13 / Small ribosomal subunit protein bS16 / Small ribosomal subunit protein bS18 / Small ribosomal subunit protein uS19 / Small ribosomal subunit protein bS20 / Small ribosomal subunit protein uS3 / Small ribosomal subunit protein uS4 / Small ribosomal subunit protein uS5 / Small ribosomal subunit protein uS8 / Small ribosomal subunit protein uS9 / Small ribosomal subunit protein uS15 / Small ribosomal subunit protein uS14 / Small ribosomal subunit protein uS17 類似検索 - 構成要素
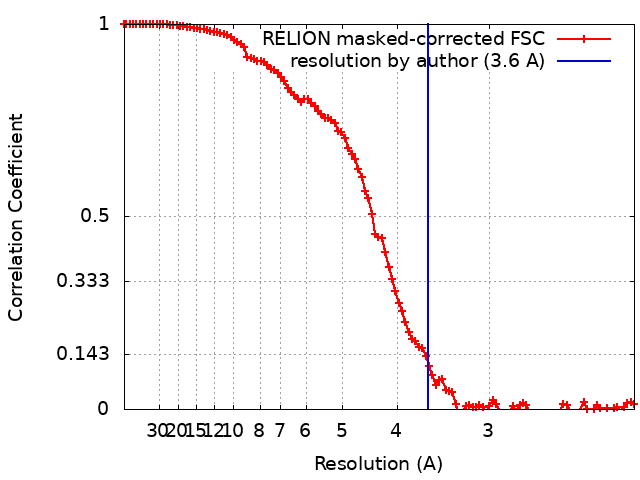
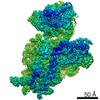
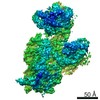
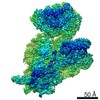
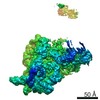
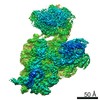
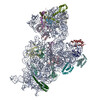
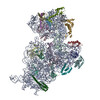
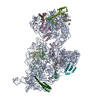
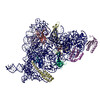
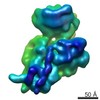
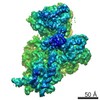
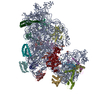
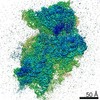
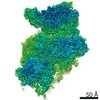
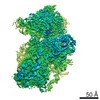
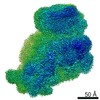
ジャーナル: RNA / 年: 2020 タイトル: Alternative conformations and motions adopted by 30S ribosomal subunits visualized by cryo-electron microscopy. 著者: Dushyant Jahagirdar / Vikash Jha / Kaustuv Basu / Josue Gomez-Blanco / Javier Vargas / Joaquin Ortega / 要旨: It is only after recent advances in cryo-electron microscopy that it is now possible to describe at high-resolution structures of large macromolecules that do not crystalize. Purified 30S subunits ...It is only after recent advances in cryo-electron microscopy that it is now possible to describe at high-resolution structures of large macromolecules that do not crystalize. Purified 30S subunits interconvert between an "active" and "inactive" conformation. The active conformation was described by crystallography in the early 2000s, but the structure of the inactive form at high resolution remains unsolved. Here we used cryo-electron microscopy to obtain the structure of the inactive conformation of the 30S subunit to 3.6 Å resolution and study its motions. In the inactive conformation, an alternative base-pairing of three nucleotides causes the region of helix 44, forming the decoding center to adopt an unlatched conformation and the 3' end of the 16S rRNA positions similarly to the mRNA during translation. Incubation of inactive 30S subunits at 42°C reverts these structural changes. The air-water interface to which ribosome subunits are exposed during sample preparation also peel off some ribosomal proteins. Extended exposures to low magnesium concentrations make the ribosomal particles more susceptible to the air-water interface causing the unfolding of large rRNA structural domains. Overall, this study provides new insights about the conformational space explored by the 30S ribosomal subunit when the ribosomal particles are free in solution.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 カナダ, 1件
カナダ, 1件  引用
引用 構造の表示
構造の表示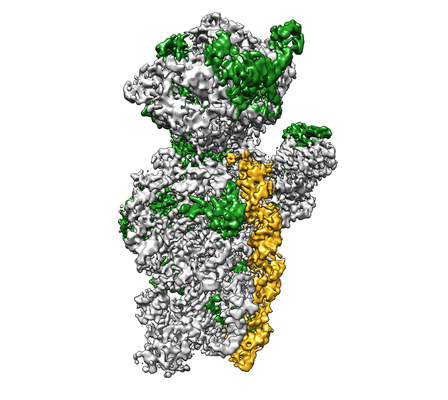
 ダウンロードとリンク
ダウンロードとリンク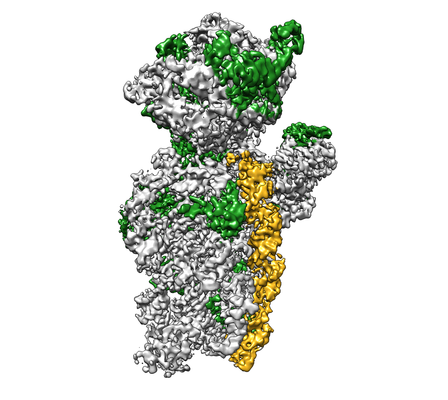 emd_21558.png
emd_21558.png http://ftp.pdbj.org/pub/emdb/structures/EMD-21558
http://ftp.pdbj.org/pub/emdb/structures/EMD-21558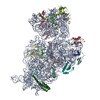







 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)




















 試料の構成要素
試料の構成要素 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN