 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 |  | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
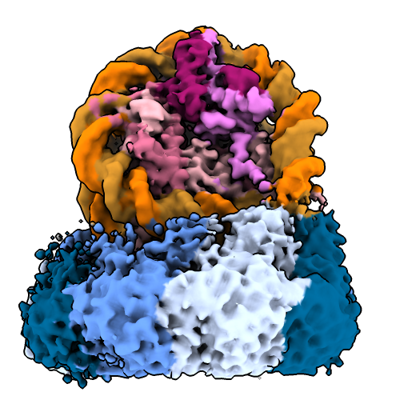
| タイトル | The cryo-EM structure of the nonameric RAD51 ring bound to the nucleosome with the linker DNA binding | ||||||||||||||||||
 マップデータ マップデータ | |||||||||||||||||||
 試料 試料 |
| ||||||||||||||||||
 キーワード キーワード |  Nucleosome (ヌクレオソーム) / Recombinase / DNA BINDING PROTEIN-DNA Complex Nucleosome (ヌクレオソーム) / Recombinase / DNA BINDING PROTEIN-DNA Complex | ||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報presynaptic intermediate filament cytoskeleton / mitotic recombination-dependent replication fork processing / chromosome organization involved in meiotic cell cycle / cellular response to camptothecin / DNA recombinase assembly / telomere maintenance via telomere lengthening / positive regulation of DNA ligation / double-strand break repair involved in meiotic recombination / nuclear ubiquitin ligase complex / mitotic recombination ...presynaptic intermediate filament cytoskeleton / mitotic recombination-dependent replication fork processing / chromosome organization involved in meiotic cell cycle / cellular response to camptothecin / DNA recombinase assembly / telomere maintenance via telomere lengthening / positive regulation of DNA ligation / double-strand break repair involved in meiotic recombination / nuclear ubiquitin ligase complex / mitotic recombination / DNA strand invasion / cellular response to hydroxyurea / replication-born double-strand break repair via sister chromatid exchange / DNA strand exchange activity / lateral element / telomere maintenance via recombination / regulation of DNA damage checkpoint / Impaired BRCA2 binding to PALB2 / single-stranded DNA helicase activity / reciprocal meiotic recombination / Defective homologous recombination repair (HRR) due to BRCA1 loss of function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA1 binding function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA2/RAD51/RAD51C binding function / Homologous DNA Pairing and Strand Exchange / Resolution of D-loop Structures through Synthesis-Dependent Strand Annealing (SDSA) / Resolution of D-loop Structures through Holliday Junction Intermediates / HDR through Single Strand Annealing (SSA) / ATP-dependent DNA damage sensor activity / Impaired BRCA2 binding to RAD51 / regulation of double-strand break repair via homologous recombination / nuclear chromosome / DNA unwinding involved in DNA replication / replication fork processing / Transcriptional Regulation by E2F6 / Presynaptic phase of homologous DNA pairing and strand exchange / negative regulation of tumor necrosis factor-mediated signaling pathway / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / ATP-dependent activity, acting on DNA / interstrand cross-link repair / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / epigenetic regulation of gene expression / DNA polymerase binding / Packaging Of Telomere Ends / condensed chromosome / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / Inhibition of DNA recombination at telomere / Meiotic synapsis / telomere organization / RNA Polymerase I Promoter Opening / Interleukin-7 signaling / Assembly of the ORC complex at the origin of replication / SUMOylation of chromatin organization proteins / DNAメチル化 / Condensation of Prophase Chromosomes / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / meiotic cell cycle / HCMV Late Events / innate immune response in mucosa / PRC2 methylates histones and DNA / condensed nuclear chromosome / male germ cell nucleus / Defective pyroptosis / cellular response to ionizing radiation / HDACs deacetylate histones / RNA Polymerase I Promoter Escape / lipopolysaccharide binding / Nonhomologous End-Joining (NHEJ) / Transcriptional regulation by small RNAs / double-strand break repair via homologous recombination / regulation of protein phosphorylation / Formation of the beta-catenin:TCF transactivating complex / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / NoRC negatively regulates rRNA expression / HDR through Homologous Recombination (HRR) / B-WICH complex positively regulates rRNA expression / G2/M DNA damage checkpoint / HDMs demethylate histones / DNA Damage/Telomere Stress Induced Senescence / PML body / Metalloprotease DUBs / PKMTs methylate histone lysines / RMTs methylate histone arginines / 遺伝的組換え / Pre-NOTCH Transcription and Translation / nucleosome assembly / Activation of anterior HOX genes in hindbrain development during early embryogenesis / HCMV Early Events / Transcriptional regulation of granulopoiesis / structural constituent of chromatin / UCH proteinases類似検索 - 分子機能 | ||||||||||||||||||
| 生物種 |  Homo sapiens (ヒト) / synthetic construct (人工物) Homo sapiens (ヒト) / synthetic construct (人工物) | ||||||||||||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.66 Å | ||||||||||||||||||
 データ登録者 データ登録者 | Shioi T / Hatazawa S / Ogasawara M / Takizawa Y / Kurumizaka H | ||||||||||||||||||
| 資金援助 |  日本, 5件 日本, 5件
| ||||||||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2024 タイトル: Cryo-EM structures of RAD51 assembled on nucleosomes containing a DSB site. 著者: Takuro Shioi / Suguru Hatazawa / Eriko Oya / Noriko Hosoya / Wataru Kobayashi / Mitsuo Ogasawara / Takehiko Kobayashi / Yoshimasa Takizawa / Hitoshi Kurumizaka / 要旨: RAD51 is the central eukaryotic recombinase required for meiotic recombination and mitotic repair of double-strand DNA breaks (DSBs). However, the mechanism by which RAD51 functions at DSB sites in ...RAD51 is the central eukaryotic recombinase required for meiotic recombination and mitotic repair of double-strand DNA breaks (DSBs). However, the mechanism by which RAD51 functions at DSB sites in chromatin has remained elusive. Here we report the cryo-electron microscopy structures of human RAD51-nucleosome complexes, in which RAD51 forms ring and filament conformations. In the ring forms, the N-terminal lobe domains (NLDs) of RAD51 protomers are aligned on the outside of the RAD51 ring, and directly bind to the nucleosomal DNA. The nucleosomal linker DNA that contains the DSB site is recognized by the L1 and L2 loops-active centres that face the central hole of the RAD51 ring. In the filament form, the nucleosomal DNA is peeled by the RAD51 filament extension, and the NLDs of RAD51 protomers proximal to the nucleosome bind to the remaining nucleosomal DNA and histones. Mutations that affect nucleosome-binding residues of the RAD51 NLD decrease nucleosome binding, but barely affect DNA binding in vitro. Consistently, yeast Rad51 mutants with the corresponding mutations are substantially defective in DNA repair in vivo. These results reveal an unexpected function of the RAD51 NLD, and explain the mechanism by which RAD51 associates with nucleosomes, recognizes DSBs and forms the active filament in chromatin. | ||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
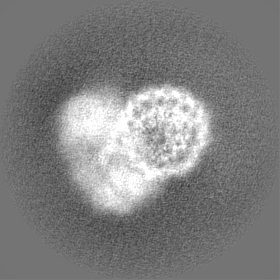
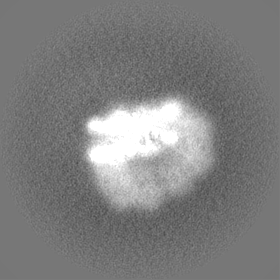
| 添付画像 |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_36442.map.gz | 9.6 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-36442-v30.xmlemd-36442.xml | 21.7 KB 21.7 KB | 表示 表示 | EMDBヘッダ |
| FSC (解像度算出) | emd_36442_fsc.xml | 10 KB | 表示 | FSCデータファイル |
| 画像 |  emd_36442.png emd_36442.png | 172.1 KB | ||
| Filedesc metadata | emd-36442.cif.gz | 6.6 KB | ||
| その他 | emd_36442_half_map_1.map.gzemd_36442_half_map_2.map.gz | 65.4 MB 65.4 MB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-36442ftp://ftp.pdbj.org/pub/emdb/structures/EMD-36442 http://ftp.pdbj.org/pub/emdb/structures/EMD-36442ftp://ftp.pdbj.org/pub/emdb/structures/EMD-36442 | HTTPS FTP |
-関連構造データ

















-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |

















-マップ
| ファイル | ダウンロード / ファイル: emd_36442.map.gz / 形式: CCP4 / 大きさ: 83.7 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ボクセルのサイズ | X=Y=Z: 1.06 Å | ||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||
| 詳細 | EMDB XML:
|
-添付データ
-ハーフマップ: #2
| ファイル | emd_36442_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |
 Z
Z Y
Y X
X

-ハーフマップ: #1
| ファイル | emd_36442_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |

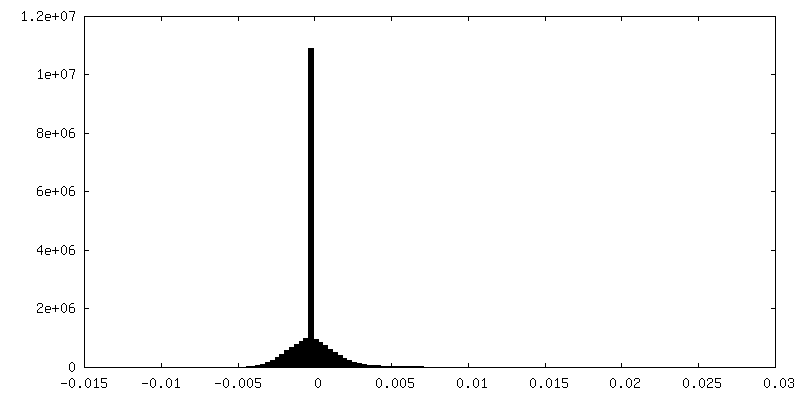
- 試料の構成要素
試料の構成要素
-全体 : RAD51-nucleosome complex
| 全体 | 名称: RAD51-nucleosome complex |
|---|---|
| 要素 |
|
-超分子 #1: RAD51-nucleosome complex
| 超分子 | 名称: RAD51-nucleosome complex / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1-#7 |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
-分子 #1: Histone H3.1
| 分子 | 名称: Histone H3.1 / タイプ: protein_or_peptide / ID: 1 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 15.719445 KDa |
| 組換発現 | 生物種:  Escherichia coli (大腸菌) Escherichia coli (大腸菌) |
| 配列 | 文字列: GSHMARTKQT ARKSTGGKAP RKQLATKAAR KSAPATGGVK KPHRYRPGTV ALREIRRYQK STELLIRKLP FQRLVREIAQ DFKTDLRFQ SSAVMALQEA CEAYLVGLFE DTNLCAIHAK RVTIMPKDIQ LARRIRGERA UniProtKB: ヒストンH3 |
-分子 #2: Histone H4
| 分子 | 名称: Histone H4 / タイプ: protein_or_peptide / ID: 2 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 11.676703 KDa |
| 組換発現 | 生物種: Escherichia coli (大腸菌) |
| 配列 | 文字列: GSHMSGRGKG GKGLGKGGAK RHRKVLRDNI QGITKPAIRR LARRGGVKRI SGLIYEETRG VLKVFLENVI RDAVTYTEHA KRKTVTAMD VVYALKRQGR TLYGFGG UniProtKB: ヒストンH4 |
-分子 #3: Histone H2A type 1-B/E
| 分子 | 名称: Histone H2A type 1-B/E / タイプ: protein_or_peptide / ID: 3 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 14.447825 KDa |
| 組換発現 | 生物種: Escherichia coli (大腸菌) |
| 配列 | 文字列: GSHMSGRGKQ GGKARAKAKT RSSRAGLQFP VGRVHRLLRK GNYSERVGAG APVYLAAVLE YLTAEILELA GNAARDNKKT RIIPRHLQL AIRNDEELNK LLGRVTIAQG GVLPNIQAVL LPKKTESHHK AKGK UniProtKB: Histone H2A type 1-B/E |
-分子 #4: Histone H2B type 1-J
| 分子 | 名称: Histone H2B type 1-J / タイプ: protein_or_peptide / ID: 4 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 14.217516 KDa |
| 組換発現 | 生物種: Escherichia coli (大腸菌) |
| 配列 | 文字列: GSHMPEPAKS APAPKKGSKK AVTKAQKKDG KKRKRSRKES YSIYVYKVLK QVHPDTGISS KAMGIMNSFV NDIFERIAGE ASRLAHYNK RSTITSREIQ TAVRLLLPGE LAKHAVSEGT KAVTKYTSAK UniProtKB: Histone H2B type 1-J |
-分子 #7: DNA repair protein RAD51 homolog 1
| 分子 | 名称: DNA repair protein RAD51 homolog 1 / タイプ: protein_or_peptide / ID: 7 / コピー数: 9 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 37.291398 KDa |
| 組換発現 | 生物種: Escherichia coli (大腸菌) |
| 配列 | 文字列: GSHMAMQMQL EANADTSVEE ESFGPQPISR LEQCGINAND VKKLEEAGFH TVEAVAYAPK KELINIKGIS EAKADKILAE AAKLVPMGF TTATEFHQRR SEIIQITTGS KELDKLLQGG IETGSITEMF GEFRTGKTQI CHTLAVTCQL PIDRGGGEGK A MYIDTEGT ...文字列: GSHMAMQMQL EANADTSVEE ESFGPQPISR LEQCGINAND VKKLEEAGFH TVEAVAYAPK KELINIKGIS EAKADKILAE AAKLVPMGF TTATEFHQRR SEIIQITTGS KELDKLLQGG IETGSITEMF GEFRTGKTQI CHTLAVTCQL PIDRGGGEGK A MYIDTEGT FRPERLLAVA ERYGLSGSDV LDNVAYARAF NTDHQTQLLY QASAMMVESR YALLIVDSAT ALYRTDYSGR GE LSARQMH LARFLRMLLR LADEFGVAVV ITNQVVAQVD GAAMFAADPK KPIGGNIIAH ASTTRLYLRK GRGETRICKI YDS PCLPEA EAMFAINADG VGDAKD UniProtKB: DNA repair protein RAD51 homolog 1 |
-分子 #5: DNA (156-MER)
| 分子 | 名称: DNA (156-MER) / タイプ: dna / ID: 5 / コピー数: 1 / 分類: DNA |
|---|---|
| 由来(天然) | 生物種: synthetic construct (人工物) |
| 分子量 | 理論値: 47.976699 KDa |
| 配列 | 文字列: (DA)(DT)(DC)(DA)(DG)(DA)(DA)(DT)(DC)(DC) (DC)(DG)(DG)(DT)(DG)(DC)(DC)(DG)(DA)(DG) (DG)(DC)(DC)(DG)(DC)(DT)(DC)(DA)(DA) (DT)(DT)(DG)(DG)(DT)(DC)(DG)(DT)(DA)(DG) (DA) (DC)(DA)(DG)(DC)(DT) ...文字列: (DA)(DT)(DC)(DA)(DG)(DA)(DA)(DT)(DC)(DC) (DC)(DG)(DG)(DT)(DG)(DC)(DC)(DG)(DA)(DG) (DG)(DC)(DC)(DG)(DC)(DT)(DC)(DA)(DA) (DT)(DT)(DG)(DG)(DT)(DC)(DG)(DT)(DA)(DG) (DA) (DC)(DA)(DG)(DC)(DT)(DC)(DT)(DA) (DG)(DC)(DA)(DC)(DC)(DG)(DC)(DT)(DT)(DA) (DA)(DA) (DC)(DG)(DC)(DA)(DC)(DG)(DT) (DA)(DC)(DG)(DC)(DG)(DC)(DT)(DG)(DT)(DC) (DC)(DC)(DC) (DC)(DG)(DC)(DG)(DT)(DT) (DT)(DT)(DA)(DA)(DC)(DC)(DG)(DC)(DC)(DA) (DA)(DG)(DG)(DG) (DG)(DA)(DT)(DT)(DA) (DC)(DA)(DC)(DC)(DC)(DA)(DA)(DG)(DA)(DC) (DA)(DC)(DC)(DA)(DG) (DG)(DC)(DA)(DC) (DG)(DA)(DG)(DA)(DC)(DA)(DG)(DA)(DA)(DA) (DA)(DA)(DA)(DA)(DC)(DA) (DA)(DC)(DG) (DA)(DA)(DA)(DA)(DC)(DG)(DG)(DC)(DC)(DA) (DC)(DC)(DA) |
-分子 #6: DNA (153-MER)
| 分子 | 名称: DNA (153-MER) / タイプ: dna / ID: 6 / コピー数: 1 / 分類: DNA |
|---|---|
| 由来(天然) | 生物種: synthetic construct (人工物) |
| 分子量 | 理論値: 47.37107 KDa |
| 配列 | 文字列: (DT)(DG)(DG)(DC)(DC)(DG)(DT)(DT)(DT)(DT) (DC)(DG)(DT)(DT)(DG)(DT)(DT)(DT)(DT)(DT) (DT)(DT)(DC)(DT)(DG)(DT)(DC)(DT)(DC) (DG)(DT)(DG)(DC)(DC)(DT)(DG)(DG)(DT)(DG) (DT) (DC)(DT)(DT)(DG)(DG) ...文字列: (DT)(DG)(DG)(DC)(DC)(DG)(DT)(DT)(DT)(DT) (DC)(DG)(DT)(DT)(DG)(DT)(DT)(DT)(DT)(DT) (DT)(DT)(DC)(DT)(DG)(DT)(DC)(DT)(DC) (DG)(DT)(DG)(DC)(DC)(DT)(DG)(DG)(DT)(DG) (DT) (DC)(DT)(DT)(DG)(DG)(DG)(DT)(DG) (DT)(DA)(DA)(DT)(DC)(DC)(DC)(DC)(DT)(DT) (DG)(DG) (DC)(DG)(DG)(DT)(DT)(DA)(DA) (DA)(DA)(DC)(DG)(DC)(DG)(DG)(DG)(DG)(DG) (DA)(DC)(DA) (DG)(DC)(DG)(DC)(DG)(DT) (DA)(DC)(DG)(DT)(DG)(DC)(DG)(DT)(DT)(DT) (DA)(DA)(DG)(DC) (DG)(DG)(DT)(DG)(DC) (DT)(DA)(DG)(DA)(DG)(DC)(DT)(DG)(DT)(DC) (DT)(DA)(DC)(DG)(DA) (DC)(DC)(DA)(DA) (DT)(DT)(DG)(DA)(DG)(DC)(DG)(DG)(DC)(DC) (DT)(DC)(DG)(DG)(DC)(DA) (DC)(DC)(DG) (DG)(DG)(DA)(DT)(DT)(DC)(DT)(DG)(DA)(DT) |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 緩衝液 | pH: 7.5 |
|---|---|
| 凍結 | 凍結剤: ETHANE |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 電子線 | 加速電圧: 300 kV / 電子線源: FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELDBright-field microscopy / 最大 デフォーカス(公称値): 2.5 µm / 最小 デフォーカス(公称値): 1.0 µm |
| 撮影 | フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 平均電子線量: 60.0 e/Å2 |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
-画像解析
| 初期モデル | モデルのタイプ: PDB ENTRY PDBモデル - PDB ID: 詳細: 5Y0C and 7OHC were also used. |
|---|---|
| 初期 角度割当 | タイプ: MAXIMUM LIKELIHOOD |
| 最終 角度割当 | タイプ: MAXIMUM LIKELIHOOD |
| 最終 再構成 | 解像度のタイプ: BY AUTHOR / 解像度: 3.66 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 使用した粒子像数: 63530 |
| FSC曲線 (解像度の算出) | 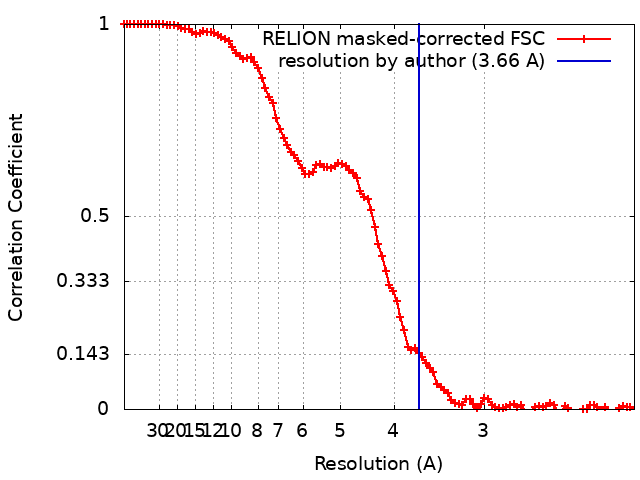 |

