 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 9i3i | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
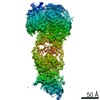
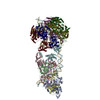
| タイトル | Cryo-EM structure of the MCM-ORC (MO) complex featuring an ORC2 regulatory domain involved in cell cycle regulation of MCM-DH loading for DNA replication. | |||||||||||||||||||||
 要素 要素 |
| |||||||||||||||||||||
 キーワード キーワード | REPLICATION / DNA Replication / Origin licensing / MCM2-7 helicase / Origin Recognition Complex / CDK / cell cycle | |||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報CDC6 association with the ORC:origin complex / Cul8-RING ubiquitin ligase complex / maintenance of rDNA / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / MCM complex binding / nuclear DNA replication / premeiotic DNA replication / Assembly of the ORC complex at the origin of replication ...CDC6 association with the ORC:origin complex / Cul8-RING ubiquitin ligase complex / maintenance of rDNA / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / MCM complex binding / nuclear DNA replication / premeiotic DNA replication / Assembly of the ORC complex at the origin of replication / replication fork protection complex / nuclear origin of replication recognition complex / pre-replicative complex assembly involved in nuclear cell cycle DNA replication / mitotic DNA replication / Activation of the pre-replicative complex / CMG complex / nuclear pre-replicative complex / nucleosome organization / Activation of ATR in response to replication stress / DNA replication preinitiation complex / MCM complex / mitotic DNA replication checkpoint signaling / double-strand break repair via break-induced replication / mitotic DNA replication initiation / single-stranded DNA helicase activity / regulation of DNA-templated DNA replication initiation / silent mating-type cassette heterochromatin formation / DNA strand elongation involved in DNA replication / Orc1 removal from chromatin / nuclear replication fork / regulation of DNA replication / DNA replication origin binding / DNA replication initiation / subtelomeric heterochromatin formation / nucleosome binding / DNA helicase activity / transcription elongation by RNA polymerase II / helicase activity / heterochromatin formation / single-stranded DNA binding / DNA helicase / chromosome, telomeric region / DNA replication / DNA damage response / chromatin binding / ATP hydrolysis activity / zinc ion binding / nucleoplasm / ATP binding / metal ion binding / nucleus / cytoplasm 類似検索 - 分子機能 | |||||||||||||||||||||
| 生物種 |  | |||||||||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.4 Å | |||||||||||||||||||||
 データ登録者 データ登録者 | Miller, T.C.R. / Lim, C.T. / Diffley, J.F.X. / Costa, A. | |||||||||||||||||||||
| 資金援助 |  英国, European Union, 英国, European Union,  デンマーク, 6件 デンマーク, 6件
| |||||||||||||||||||||
 引用 引用 | ジャーナル: Nat Struct Mol Biol / 年: 2025 タイトル: Cell cycle regulation has shaped replication origins in budding yeast. 著者: Chew Theng Lim / Thomas C R Miller / Kang Wei Tan / Saurabh Talele / Anne Early / Philip East / Humberto Sánchez / Nynke H Dekker / Alessandro Costa / John F X Diffley /  要旨: Eukaryotic DNA replication initiates from genomic loci known as origins. At budding yeast origins like ARS1, a double hexamer (DH) of the MCM replicative helicase is assembled by origin recognition ...Eukaryotic DNA replication initiates from genomic loci known as origins. At budding yeast origins like ARS1, a double hexamer (DH) of the MCM replicative helicase is assembled by origin recognition complex (ORC), Cdc6 and Cdt1 by sequential hexamer loading from two opposed ORC binding sites. Cyclin-dependent kinase (CDK) inhibits DH assembly, which prevents re-replication by restricting helicase loading to the G1 phase. Here, we show that an intrinsically disordered region (IDR) in the Orc2 subunit promotes interaction between ORC and the first loaded, closed-ring MCM hexamer (the MCM-ORC (MO) intermediate). CDK-dependent phosphorylation of this IDR blocks MO formation and DH assembly. We show that MO stabilizes ORC at lower-affinity binding sites required for second hexamer loading. Origins comprising two high-affinity ORC sites can assemble DH efficiently without MO by independently loading single hexamers. Strikingly, these origins escape CDK inhibition in vitro and in vivo. Our work reveals mechanistic plasticity in MCM loading with implications for understanding how CDK regulation has shaped yeast origin evolution and how natural, strong origins might escape cell cycle regulation. We also identify key steps common to loading pathways, with implications for understanding how MCM is loaded in other eukaryotes. #1: ジャーナル: Nature / 年: 2019タイトル: Mechanism of head-to-head MCM double-hexamer formation revealed by cryo-EM. 著者: Thomas C R Miller / Julia Locke / Julia F Greiwe / John F X Diffley / Alessandro Costa / 要旨: In preparation for bidirectional DNA replication, the origin recognition complex (ORC) loads two hexameric MCM helicases to form a head-to-head double hexamer around DNA. The mechanism of MCM double- ...In preparation for bidirectional DNA replication, the origin recognition complex (ORC) loads two hexameric MCM helicases to form a head-to-head double hexamer around DNA. The mechanism of MCM double-hexamer formation is debated. Single-molecule experiments have suggested a sequential mechanism, in which the ORC-dependent loading of the first hexamer drives the recruitment of the second hexamer. By contrast, biochemical data have shown that two rings are loaded independently via the same ORC-mediated mechanism, at two inverted DNA sites. Here we visualize MCM loading using time-resolved electron microscopy, and identify intermediates in the formation of the double hexamer. We confirm that both hexamers are recruited via the same interaction that occurs between ORC and the C-terminal domains of the MCM helicases. Moreover, we identify the mechanism of coupled MCM loading. The loading of the first MCM hexamer around DNA creates a distinct interaction site, which promotes the engagement of ORC at the N-terminal homodimerization interface of MCM. In this configuration, ORC is poised to direct the recruitment of the second hexamer in an inverted orientation, which is suitable for the formation of the double hexamer. Our results therefore reconcile the two apparently contrasting models derived from single-molecule experiments and biochemical data. | |||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 9i3i.cif.gz | 2.2 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb9i3i.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 9i3i.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 9i3i_validation.pdf.gz | 1.7 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 9i3i_full_validation.pdf.gz | 1.8 MB | 表示 | |
| XML形式データ | 9i3i_validation.xml.gz | 167.2 KB | 表示 | |
| CIF形式データ | 9i3i_validation.cif.gz | 250.9 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/i3/9i3iftp://data.pdbj.org/pub/pdb/validation_reports/i3/9i3i | HTTPS FTP |
-関連構造データ




-リンク
 PDBj
PDBj









































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-Origin recognition complex subunit ... , 6種, 6分子 ABCDEF
| #1: タンパク質 | 分子量: 108612.922 Da / 分子数: 1 / 由来タイプ: 組換発現 / 詳細: N-terminus contains a CBP-TEV purification tag. 由来: (組換発現) 遺伝子: ORC1, YML065W / 発現宿主: |
|---|---|
| #2: タンパク質 | 分子量: 71342.180 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: ORC2, RRR1, SIR5, YBR060C, YBR0523 / 発現宿主: |
| #3: タンパク質 | 分子量: 72161.766 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: ORC3, OAF1, OIF1, YLL004W, L1365 / 発現宿主: |
| #4: タンパク質 | 分子量: 60772.152 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: ORC4, YPR162C, P9325.5 / 発現宿主: |
| #5: タンパク質 | 分子量: 55347.168 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: ORC5, YNL261W, N0834 / 発現宿主: |
| #6: タンパク質 | 分子量: 50369.531 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: ORC6, AAP1, YHR118C / 発現宿主: |
-DNA replication licensing factor ... , 5種, 5分子 23467
| #7: タンパク質 | 分子量: 98911.539 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM2, YBL023C, YBL0438 / 発現宿主: |
|---|---|
| #8: タンパク質 | 分子量: 111720.242 Da / 分子数: 1 / 由来タイプ: 組換発現 / 詳細: N-terminus contains a CBP-TEV purification tag 由来: (組換発現) 遺伝子: MCM3, YEL032W, SYGP-ORF23 / 発現宿主: |
| #9: タンパク質 | 分子量: 105138.375 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM4, CDC54, HCD21, YPR019W, YP9531.13 / 発現宿主: |
| #11: タンパク質 | 分子量: 113110.211 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM6, YGL201C / 発現宿主: |
| #12: タンパク質 | 分子量: 95049.875 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM7, CDC47, YBR202W, YBR1441 / 発現宿主: |
-タンパク質 , 1種, 1分子 5
| #10: タンパク質 | 分子量: 86505.734 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM5, CDC46, YLR274W, L9328.1 / 発現宿主: |
|---|
-DNA鎖 , 2種, 2分子 XY
| #13: DNA鎖 | 分子量: 27116.334 Da / 分子数: 1 / 由来タイプ: 合成 由来: (合成) |
|---|---|
| #14: DNA鎖 | 分子量: 27154.562 Da / 分子数: 1 / 由来タイプ: 合成 由来: (合成) |
-非ポリマー , 4種, 15分子 



| #15: 化合物 |  分子量: 507.181 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM 分子量: 507.181 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM#16: 化合物 |  分子量: 24.305 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION#17: 化合物 | ChemComp-ADP /  分子量: 427.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM#18: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION 分子量: 65.409 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | N |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: MCM-ORC featuring ORC2 regulatory domain / タイプ: COMPLEX / Entity ID: #1-#14 / 由来: RECOMBINANT |
|---|---|
| 由来(天然) | 生物種: |
| 由来(組換発現) | 生物種: |
| 緩衝液 | pH: 7.6 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 装置: LEICA EM GP / 凍結剤: ETHANE / 湿度: 90 % / 凍結前の試料温度: 288 K 詳細: 10 second incubation, 3.5 seconds single side blotting. |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: TFS KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 4200 nm / 最小 デフォーカス(公称値): 2700 nm |
| 試料ホルダ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 50.4 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 1 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 6287507 | ||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 4.4 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 177687 / 対称性のタイプ: POINT |
