 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-9i3i: Cryo-EM structure of the MCM-ORC (MO) complex featuring an ORC2 r... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 9i3i | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of the MCM-ORC (MO) complex featuring an ORC2 regulatory domain involved in cell cycle regulation of MCM-DH loading for DNA replication. | |||||||||||||||||||||
 Components Components |
| |||||||||||||||||||||
 Keywords Keywords | REPLICATION / DNA Replication / Origin licensing / MCM2-7 helicase / Origin Recognition Complex / CDK / cell cycle | |||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationCDC6 association with the ORC:origin complex / maintenance of rDNA / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / MCM complex binding / nuclear DNA replication / Assembly of the ORC complex at the origin of replication / premeiotic DNA replication / pre-replicative complex assembly involved in nuclear cell cycle DNA replication ...CDC6 association with the ORC:origin complex / maintenance of rDNA / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / MCM complex binding / nuclear DNA replication / Assembly of the ORC complex at the origin of replication / premeiotic DNA replication / pre-replicative complex assembly involved in nuclear cell cycle DNA replication / nuclear origin of replication recognition complex / Activation of the pre-replicative complex / nuclear pre-replicative complex / CMG complex / nucleosome organization / DNA replication preinitiation complex / Activation of ATR in response to replication stress / mitotic DNA replication checkpoint signaling / double-strand break repair via break-induced replication / MCM complex / mitotic DNA replication initiation / mitotic DNA replication / silent mating-type cassette heterochromatin formation / single-stranded DNA helicase activity / regulation of DNA-templated DNA replication initiation / Orc1 removal from chromatin / DNA strand elongation involved in DNA replication / regulation of DNA replication / nuclear replication fork / DNA replication origin binding / subtelomeric heterochromatin formation / DNA replication initiation / nucleosome binding / DNA helicase activity / helicase activity / transcription elongation by RNA polymerase II / peroxisome / single-stranded DNA binding / heterochromatin formation / DNA helicase / chromosome, telomeric region / DNA replication / chromatin binding / DNA damage response / ATP hydrolysis activity / zinc ion binding / nucleoplasm / ATP binding / metal ion binding / nucleus / cytoplasm Similarity search - Function | |||||||||||||||||||||
| Biological species |  | |||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4.4 Å | |||||||||||||||||||||
 Authors Authors | Miller, T.C.R. / Lim, C.T. / Diffley, J.F.X. / Costa, A. | |||||||||||||||||||||
| Funding support |  United Kingdom, European Union, United Kingdom, European Union,  Denmark, 6items Denmark, 6items
| |||||||||||||||||||||
 Citation Citation | Journal: Nat Struct Mol Biol / Year: 2025 Title: Cell cycle regulation has shaped replication origins in budding yeast. Authors: Chew Theng Lim / Thomas C R Miller / Kang Wei Tan / Saurabh Talele / Anne Early / Philip East / Humberto Sánchez / Nynke H Dekker / Alessandro Costa / John F X Diffley /  Abstract: Eukaryotic DNA replication initiates from genomic loci known as origins. At budding yeast origins like ARS1, a double hexamer (DH) of the MCM replicative helicase is assembled by origin recognition ...Eukaryotic DNA replication initiates from genomic loci known as origins. At budding yeast origins like ARS1, a double hexamer (DH) of the MCM replicative helicase is assembled by origin recognition complex (ORC), Cdc6 and Cdt1 by sequential hexamer loading from two opposed ORC binding sites. Cyclin-dependent kinase (CDK) inhibits DH assembly, which prevents re-replication by restricting helicase loading to the G1 phase. Here, we show that an intrinsically disordered region (IDR) in the Orc2 subunit promotes interaction between ORC and the first loaded, closed-ring MCM hexamer (the MCM-ORC (MO) intermediate). CDK-dependent phosphorylation of this IDR blocks MO formation and DH assembly. We show that MO stabilizes ORC at lower-affinity binding sites required for second hexamer loading. Origins comprising two high-affinity ORC sites can assemble DH efficiently without MO by independently loading single hexamers. Strikingly, these origins escape CDK inhibition in vitro and in vivo. Our work reveals mechanistic plasticity in MCM loading with implications for understanding how CDK regulation has shaped yeast origin evolution and how natural, strong origins might escape cell cycle regulation. We also identify key steps common to loading pathways, with implications for understanding how MCM is loaded in other eukaryotes. #1: Journal: Nature / Year: 2019Title: Mechanism of head-to-head MCM double-hexamer formation revealed by cryo-EM. Authors: Thomas C R Miller / Julia Locke / Julia F Greiwe / John F X Diffley / Alessandro Costa / Abstract: In preparation for bidirectional DNA replication, the origin recognition complex (ORC) loads two hexameric MCM helicases to form a head-to-head double hexamer around DNA. The mechanism of MCM double- ...In preparation for bidirectional DNA replication, the origin recognition complex (ORC) loads two hexameric MCM helicases to form a head-to-head double hexamer around DNA. The mechanism of MCM double-hexamer formation is debated. Single-molecule experiments have suggested a sequential mechanism, in which the ORC-dependent loading of the first hexamer drives the recruitment of the second hexamer. By contrast, biochemical data have shown that two rings are loaded independently via the same ORC-mediated mechanism, at two inverted DNA sites. Here we visualize MCM loading using time-resolved electron microscopy, and identify intermediates in the formation of the double hexamer. We confirm that both hexamers are recruited via the same interaction that occurs between ORC and the C-terminal domains of the MCM helicases. Moreover, we identify the mechanism of coupled MCM loading. The loading of the first MCM hexamer around DNA creates a distinct interaction site, which promotes the engagement of ORC at the N-terminal homodimerization interface of MCM. In this configuration, ORC is poised to direct the recruitment of the second hexamer in an inverted orientation, which is suitable for the formation of the double hexamer. Our results therefore reconcile the two apparently contrasting models derived from single-molecule experiments and biochemical data. | |||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9i3i.cif.gz | 2.2 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9i3i.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 9i3i.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/i3/9i3iftp://data.pdbj.org/pub/pdb/validation_reports/i3/9i3i | HTTPS FTP |
|---|
-Related structure data
| Related structure data | 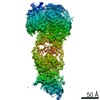 4980M  8rifC  8rigC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |


-Links
 PDBj
PDBj









































- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Origin recognition complex subunit ... , 6 types, 6 molecules ABCDEF
| #1: Protein | Mass: 108612.922 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: N-terminus contains a CBP-TEV purification tag. / Source: (gene. exp.) |
|---|---|
| #2: Protein | Mass: 71342.180 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
| #3: Protein | Mass: 72161.766 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
| #4: Protein | Mass: 60772.152 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
| #5: Protein | Mass: 55347.168 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
| #6: Protein | Mass: 50369.531 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
-DNA replication licensing factor ... , 5 types, 5 molecules 23467
| #7: Protein | Mass: 98911.539 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
|---|---|
| #8: Protein | Mass: 111720.242 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: N-terminus contains a CBP-TEV purification tag / Source: (gene. exp.) |
| #9: Protein | Mass: 105138.375 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
| #11: Protein | Mass: 113110.211 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
| #12: Protein | Mass: 95049.875 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
-Protein , 1 types, 1 molecules 5
| #10: Protein | Mass: 86505.734 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
|---|
-DNA chain , 2 types, 2 molecules XY
| #13: DNA chain | Mass: 27116.334 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) |
|---|---|
| #14: DNA chain | Mass: 27154.562 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) |
-Non-polymers , 4 types, 15 molecules 



| #15: Chemical |  Mass: 507.181 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP, energy-carrying molecule*YM#16: Chemical |  Mass: 24.305 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION Mass: 24.305 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION#17: Chemical | ChemComp-ADP /  Mass: 427.201 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Feature type: SUBJECT OF INVESTIGATION / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Feature type: SUBJECT OF INVESTIGATION / Comment: ADP, energy-carrying molecule*YM#18: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 5 / Source method: obtained synthetically / Formula: Zn / Feature type: SUBJECT OF INVESTIGATION Mass: 65.409 Da / Num. of mol.: 5 / Source method: obtained synthetically / Formula: Zn / Feature type: SUBJECT OF INVESTIGATION |
|---|
-Details
| Has ligand of interest | Y |
|---|---|
| Has protein modification | N |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: MCM-ORC featuring ORC2 regulatory domain / Type: COMPLEX / Entity ID: #1-#14 / Source: RECOMBINANT |
|---|---|
| Source (natural) | Organism: |
| Source (recombinant) | Organism: |
| Buffer solution | pH: 7.6 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Instrument: LEICA EM GP / Cryogen name: ETHANE / Humidity: 90 % / Chamber temperature: 288 K Details: 10 second incubation, 3.5 seconds single side blotting. |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: TFS KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 4200 nm / Nominal defocus min: 2700 nm |
| Specimen holder | Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Electron dose: 50.4 e/Å2 / Detector mode: COUNTING / Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Num. of grids imaged: 1 |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 6287507 | ||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4.4 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 177687 / Symmetry type: POINT |
