 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-9h6x: Cryo-EM structure of lipid-bound human myoferlin (25 mol% DOPS/5 ... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 9h6x | ||||||
|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of lipid-bound human myoferlin (25 mol% DOPS/5 mol% PI(4,5)P2 nanodisc) | ||||||
 Components Components | Myoferlin | ||||||
 Keywords Keywords | MEMBRANE PROTEIN / Ferlin / multi-C2 domain protein / tail-anchored membrane protein / vesicle docking and fusion / membrane remodeling | ||||||
| Function / homology |  Function and homology information Function and homology informationregulation of neurotransmitter secretion / plasma membrane repair / blood circulation / centriolar satellite / muscle contraction / phospholipid binding / caveola / nuclear envelope / late endosome membrane / synaptic vesicle membrane ...regulation of neurotransmitter secretion / plasma membrane repair / blood circulation / centriolar satellite / muscle contraction / phospholipid binding / caveola / nuclear envelope / late endosome membrane / synaptic vesicle membrane / nuclear membrane / cytoplasmic vesicle / calcium ion binding / extracellular exosome / plasma membrane Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 2.56 Å | ||||||
 Authors Authors | Cretu, C. / Moser, T. / Preobraschenski, J. | ||||||
| Funding support |  Germany, 1items Germany, 1items
| ||||||
 Citation Citation | Journal: EMBO J / Year: 2025 Title: Structural insights into lipid membrane binding by human ferlins. Authors: Constantin Cretu / Aleksandar Chernev / Csaba Zoltán Kibédi Szabó / Vladimir Pena / Henning Urlaub / Tobias Moser / Julia Preobraschenski /  Abstract: Ferlins are ancient membrane proteins with a unique architecture, and play central roles in crucial processes that involve Ca-dependent vesicle fusion. Despite their links to multiple human diseases ...Ferlins are ancient membrane proteins with a unique architecture, and play central roles in crucial processes that involve Ca-dependent vesicle fusion. Despite their links to multiple human diseases and numerous functional studies, a mechanistic understanding of how these multi-C domain-containing proteins interact with lipid membranes to promote membrane remodelling and fusion is currently lacking. Here we obtain near-complete cryo-electron microscopy structures of human myoferlin and dysferlin in their Ca- and lipid-bound states. We show that ferlins adopt compact, ring-like tertiary structures upon membrane binding. The top arch of the ferlin ring, composed of the CC-CD region, is rigid and exhibits only little variability across the observed functional states. In contrast, the N-terminal CB and the C-terminal CF-CG domains cycle between alternative conformations and, in response to Ca, close the ferlin ring, promoting tight interaction with the target membrane. Probing key domain interfaces validates the observed architecture, and informs a model of how ferlins engage lipid bilayers in a Ca-dependent manner. This work reveals the general principles of human ferlin structures and provides a framework for future analyses of ferlin-dependent cellular functions and disease mechanisms. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9h6x.cif.gz | 452.2 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9h6x.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 9h6x.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/h6/9h6xftp://data.pdbj.org/pub/pdb/validation_reports/h6/9h6x | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  51902MC  9qkvC  9qleC  9qlfC  9qlnC  9qlsC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |





-Links
 PDBj
PDBj


- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 232625.953 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: Fer-1-like protein 3, FER1L3, MYOF / Source: (gene. exp.) Homo sapiens (human) / Gene: MYOF, FER1L3, KIAA1207 / Production host:   Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9NZM1 Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9NZM1 | ||||||
|---|---|---|---|---|---|---|---|
| #2: Chemical | 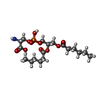  Mass: 455.437 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C18H34NO10P / Feature type: SUBJECT OF INVESTIGATION Mass: 455.437 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C18H34NO10P / Feature type: SUBJECT OF INVESTIGATION#3: Chemical | ChemComp-CA /   Mass: 40.078 Da / Num. of mol.: 10 / Source method: obtained synthetically / Formula: Ca / Feature type: SUBJECT OF INVESTIGATION Mass: 40.078 Da / Num. of mol.: 10 / Source method: obtained synthetically / Formula: Ca / Feature type: SUBJECT OF INVESTIGATIONHas ligand of interest | Y | Has protein modification | N | |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Soluble human myoferlin (residues 1-1997) bound to an MSP2N2-based lipid nanodisc (comprising 25 mol% DOPS and 5 mol% PI(4,5)P2) Type: COMPLEX / Entity ID: #1 / Source: RECOMBINANT | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Experimental value: NO | ||||||||||||||||||||||||||||||
| Source (natural) | Organism: Homo sapiens (human) | ||||||||||||||||||||||||||||||
| Source (recombinant) | Organism: Spodoptera frugiperda (fall armyworm) | ||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.5 | ||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||
| Specimen | Conc.: 0.75 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES Details: The myoferlin-nanodisc complex was assembled in vitro, crosslinked with 0.07% (v/v) glutaraldehyde, and purified by size-exclusion chromatography on Superose 6 10/300 prior to cryo-EM grid preparation | ||||||||||||||||||||||||||||||
| Specimen support | Details: Plasma Cleaner model PDC-32G-2 (Harrick Plasma) / Grid material: COPPER / Grid mesh size: 200 divisions/in. / Grid type: Quantifoil R1.2/1.3 | ||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE-PROPANE / Humidity: 100 % / Chamber temperature: 277.15 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: TFS KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 165000 X / Nominal defocus max: 2300 nm / Nominal defocus min: 800 nm / Cs: 2.7 mm / C2 aperture diameter: 50 µm / Alignment procedure: COMA FREE |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 3.16 sec. / Electron dose: 39.595 e/Å2 / Film or detector model: FEI FALCON IV (4k x 4k) / Num. of grids imaged: 1 / Num. of real images: 31039 Details: dataset 1: 8660 movies, 39.595 e-/A2 dataset 2: 22379 movies, 39.216 e-/A2 |
| EM imaging optics | Energyfilter name: TFS Selectris / Energyfilter slit width: 10 eV |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 7651843 | ||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 2.56 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 233637 / Num. of class averages: 1 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||
| Atomic model building | B value: 152.37 / Protocol: OTHER / Space: REAL Details: Iterative model building and refinement with ISOLDE, Coot, and Phenix (phenix.real_space_refine) | ||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Source name: AlphaFold / Type: in silico model | ||||||||||||||||||||||||||||||||||||||||
| Refinement | Cross valid method: NONE Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 152.4 Å2 | ||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|
