 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 9fjm | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Cryo-EM structure of the phalloidin-bound pointed end of the actin filament. | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | STRUCTURAL PROTEIN / actin / phalloidin / filament / pointed end | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報positive regulation of norepinephrine uptake / cellular response to cytochalasin B / bBAF complex / npBAF complex / nBAF complex / brahma complex / regulation of transepithelial transport / Formation of annular gap junctions / morphogenesis of a polarized epithelium / Formation of the dystrophin-glycoprotein complex (DGC) ...positive regulation of norepinephrine uptake / cellular response to cytochalasin B / bBAF complex / npBAF complex / nBAF complex / brahma complex / regulation of transepithelial transport / Formation of annular gap junctions / morphogenesis of a polarized epithelium / Formation of the dystrophin-glycoprotein complex (DGC) / structural constituent of postsynaptic actin cytoskeleton / GBAF complex / Gap junction degradation / Folding of actin by CCT/TriC / regulation of G0 to G1 transition / protein localization to adherens junction / Cell-extracellular matrix interactions / dense body / postsynaptic actin cytoskeleton / Tat protein binding / Prefoldin mediated transfer of substrate to CCT/TriC / RSC-type complex / regulation of double-strand break repair / regulation of nucleotide-excision repair / Adherens junctions interactions / RHOF GTPase cycle / adherens junction assembly / apical protein localization / Sensory processing of sound by outer hair cells of the cochlea / Interaction between L1 and Ankyrins / tight junction / SWI/SNF complex / regulation of mitotic metaphase/anaphase transition / Sensory processing of sound by inner hair cells of the cochlea / positive regulation of T cell differentiation / apical junction complex / regulation of norepinephrine uptake / positive regulation of double-strand break repair / transporter regulator activity / maintenance of blood-brain barrier / nitric-oxide synthase binding / cortical cytoskeleton / NuA4 histone acetyltransferase complex / establishment or maintenance of cell polarity / positive regulation of stem cell population maintenance / Regulation of MITF-M-dependent genes involved in pigmentation / Recycling pathway of L1 / brush border / regulation of G1/S transition of mitotic cell cycle / kinesin binding / EPH-ephrin mediated repulsion of cells / negative regulation of cell differentiation / RHO GTPases Activate WASPs and WAVEs / regulation of synaptic vesicle endocytosis / positive regulation of myoblast differentiation / RHO GTPases activate IQGAPs / regulation of protein localization to plasma membrane / positive regulation of double-strand break repair via homologous recombination / EPHB-mediated forward signaling / cytoskeleton organization / substantia nigra development / axonogenesis / calyx of Held / nitric-oxide synthase regulator activity / Translocation of SLC2A4 (GLUT4) to the plasma membrane / FCGR3A-mediated phagocytosis / actin filament / adherens junction / positive regulation of cell differentiation / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / cell motility / RHO GTPases Activate Formins / Signaling by high-kinase activity BRAF mutants / MAP2K and MAPK activation / DNA Damage Recognition in GG-NER / kinetochore / Regulation of actin dynamics for phagocytic cup formation / B-WICH complex positively regulates rRNA expression / structural constituent of cytoskeleton / 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 / platelet aggregation / VEGFA-VEGFR2 Pathway / tau protein binding / Schaffer collateral - CA1 synapse / nuclear matrix / cytoplasmic ribonucleoprotein granule / Signaling by RAF1 mutants / Signaling by moderate kinase activity BRAF mutants / Paradoxical activation of RAF signaling by kinase inactive BRAF / Signaling downstream of RAS mutants / cell-cell junction / UCH proteinases / Signaling by BRAF and RAF1 fusions / nucleosome / presynapse / lamellipodium / actin cytoskeleton / Clathrin-mediated endocytosis / HATs acetylate histones / Factors involved in megakaryocyte development and platelet production 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) Amanita phalloides (タマゴテングタケ) Amanita phalloides (タマゴテングタケ) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.65 Å | ||||||||||||
 データ登録者 データ登録者 | Boiero Sanders, M. / Oosterheert, W. / Hofnagel, O. / Bieling, P. / Raunser, S. | ||||||||||||
| 資金援助 |  ドイツ, European Union, 3件 ドイツ, European Union, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2024 タイトル: Phalloidin and DNase I-bound F-actin pointed end structures reveal principles of filament stabilization and disassembly. 著者: Micaela Boiero Sanders / Wout Oosterheert / Oliver Hofnagel / Peter Bieling / Stefan Raunser / 要旨: Actin filament turnover involves subunits binding to and dissociating from the filament ends, with the pointed end being the primary site of filament disassembly. Several molecules modulate filament ...Actin filament turnover involves subunits binding to and dissociating from the filament ends, with the pointed end being the primary site of filament disassembly. Several molecules modulate filament turnover, but the underlying mechanisms remain incompletely understood. Here, we present three cryo-EM structures of the F-actin pointed end in the presence and absence of phalloidin or DNase I. The two terminal subunits at the undecorated pointed end adopt a twisted conformation. Phalloidin can still bind and bridge these subunits, inducing a conformational shift to a flattened, F-actin-like state. This explains how phalloidin prevents depolymerization at the pointed end. Interestingly, two DNase I molecules simultaneously bind to the phalloidin-stabilized pointed end. In the absence of phalloidin, DNase I binding would disrupt the terminal actin subunit packing, resulting in filament disassembly. Our findings uncover molecular principles of pointed end regulation and provide structural insights into the kinetic asymmetry between the actin filament ends. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 9fjm.cif.gz | 265.3 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb9fjm.ent.gz | 215.7 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 9fjm.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 9fjm_validation.pdf.gz | 1.8 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 9fjm_full_validation.pdf.gz | 1.8 MB | 表示 | |
| XML形式データ | 9fjm_validation.xml.gz | 65.1 KB | 表示 | |
| CIF形式データ | 9fjm_validation.cif.gz | 92.7 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/fj/9fjmftp://data.pdbj.org/pub/pdb/validation_reports/fj/9fjm | HTTPS FTP |
-関連構造データ




-リンク
 PDBj
PDBj
















- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 41632.422 Da / 分子数: 4 / 由来タイプ: 組換発現 詳細: Beta-actin recombinantly purified from insect cells. 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: ACTB / プラスミド: p2336 pFL_ACTB_C272A / 細胞株 (発現宿主): BTI-Tnao38 / 発現宿主:  Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: P60709 Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: P60709#2: タンパク質・ペプチド | タイプ: Peptide-like / クラス: 毒素 / 分子量: 808.899 Da / 分子数: 4 / 由来タイプ: 天然 / 詳細: phalloidin from Amanita phalloides. 由来: (天然) Amanita phalloides (タマゴテングタケ)参照: BIRD: PRD_002366 #3: 化合物 | ChemComp-ADP /   分子量: 427.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM#4: 化合物 | ChemComp-MG /   分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION#5: 化合物 | ChemComp-PO4 / 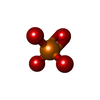  分子量: 94.971 Da / 分子数: 4 / 由来タイプ: 合成 / 式: PO4 / タイプ: SUBJECT OF INVESTIGATION 分子量: 94.971 Da / 分子数: 4 / 由来タイプ: 合成 / 式: PO4 / タイプ: SUBJECT OF INVESTIGATION研究の焦点であるリガンドがあるか | Y | |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 |
| ||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||
| 由来(組換発現) | 生物種: Trichoplusia ni (イラクサキンウワバ) / 細胞: BTI-Tnao38 | ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.1 詳細: 1xKMEH (10 mM HEPES pH 7.1, 100 mM KCl, 2 mM MgCl2, 1 mM EGTA, 0.5 mM TCEP) | ||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: GOLD / グリッドのサイズ: 200 divisions/in. / グリッドのタイプ: Quantifoil R2/1 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE-PROPANE / 湿度: 100 % / 凍結前の試料温度: 286 K / 詳細: 3 seconds, force 0. |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS 詳細: 300 kV Titan Krios G2 microscope (Thermo Fisher Scientific) with an in-column Cs-corrector. |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 81000 X / 最大 デフォーカス(公称値): 2700 nm / 最小 デフォーカス(公称値): 1200 nm / Cs: 0.01 mm / C2レンズ絞り径: 50 µm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 64.6 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 撮影したグリッド数: 1 / 実像数: 20393 |
| 電子光学装置 | エネルギーフィルター名称: GIF Bioquantum / 詳細: Gatan energy filter. / エネルギーフィルタースリット幅: 15 eV 球面収差補正装置: Titan Krios G2 microscope (Thermo Fisher Scientific) with an in-column Cs-corrector. |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 5200600 / 詳細: Particles picked using SPHIRE-crYOLO. | ||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.65 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 280802 / 詳細: Local Refinement in CryoSPARC. / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: REAL | ||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | PDB-ID: 8rtt Accession code: 8rtt / 詳細: actin and phalloidin model / Source name: PDB / タイプ: experimental model | ||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|

