 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-9fjm: Cryo-EM structure of the phalloidin-bound pointed end of the acti... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 9fjm | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of the phalloidin-bound pointed end of the actin filament. | ||||||||||||
 Components Components |
| ||||||||||||
 Keywords Keywords | STRUCTURAL PROTEIN / actin / phalloidin / filament / pointed end | ||||||||||||
| Function / homology |  Function and homology information Function and homology informationpositive regulation of norepinephrine uptake / bBAF complex / GBAF complex / brahma complex / cellular response to cytochalasin B / Formation of the embryonic stem cell BAF (esBAF) complex / npBAF complex / nBAF complex / Formation of the canonical BAF (cBAF) complex / regulation of transepithelial transport ...positive regulation of norepinephrine uptake / bBAF complex / GBAF complex / brahma complex / cellular response to cytochalasin B / Formation of the embryonic stem cell BAF (esBAF) complex / npBAF complex / nBAF complex / Formation of the canonical BAF (cBAF) complex / regulation of transepithelial transport / morphogenesis of a polarized epithelium / structural constituent of postsynaptic actin cytoskeleton / Formation of annular gap junctions / Formation of the dystrophin-glycoprotein complex (DGC) / Formation of the polybromo-BAF (pBAF) complex / Formation of neuronal progenitor and neuronal BAF (npBAF and nBAF) / Gap junction degradation / protein localization to adherens junction / Formation of the non-canonical BAF (ncBAF) complex / regulation of G0 to G1 transition / Cell-extracellular matrix interactions / dense body / Folding of actin by CCT/TriC / Tat protein binding / RSC-type complex / postsynaptic actin cytoskeleton / Regulation of CDH1 Function / regulation of double-strand break repair / apical protein localization / Prefoldin mediated transfer of substrate to CCT/TriC / Adherens junctions interactions / adherens junction assembly / RHOF GTPase cycle / regulation of nucleotide-excision repair / Sensory processing of sound by outer hair cells of the cochlea / tight junction / regulation of mitotic metaphase/anaphase transition / SWI/SNF complex / Sensory processing of sound by inner hair cells of the cochlea / Interaction between L1 and Ankyrins / positive regulation of T cell differentiation / apical junction complex / maintenance of blood-brain barrier / positive regulation of stem cell population maintenance / regulation of norepinephrine uptake / transporter regulator activity / NuA4 histone acetyltransferase complex / positive regulation of double-strand break repair / Recycling pathway of L1 / cortical cytoskeleton / Regulation of MITF-M-dependent genes involved in pigmentation / establishment or maintenance of cell polarity / nitric-oxide synthase binding / brush border / EPH-ephrin mediated repulsion of cells / negative regulation of cell differentiation / regulation of synaptic vesicle endocytosis / positive regulation of myoblast differentiation / RHO GTPases Activate WASPs and WAVEs / kinesin binding / regulation of protein localization to plasma membrane / RHO GTPases activate IQGAPs / positive regulation of double-strand break repair via homologous recombination / regulation of G1/S transition of mitotic cell cycle / axonogenesis / cytoskeleton organization / EPHB-mediated forward signaling / substantia nigra development / calyx of Held / nitric-oxide synthase regulator activity / cell motility / FCGR3A-mediated phagocytosis / Translocation of SLC2A4 (GLUT4) to the plasma membrane / actin filament / adherens junction / positive regulation of cell differentiation / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / RHO GTPases Activate Formins / Signaling by high-kinase activity BRAF mutants / MAP2K and MAPK activation / Regulation of actin dynamics for phagocytic cup formation / VEGFA-VEGFR2 Pathway / platelet aggregation / B-WICH complex positively regulates rRNA expression / tau protein binding / DNA Damage Recognition in GG-NER / Hydrolases; Acting on acid anhydrides; Acting on acid anhydrides to facilitate cellular and subcellular movement / Schaffer collateral - CA1 synapse / structural constituent of cytoskeleton / kinetochore / nuclear matrix / cytoplasmic ribonucleoprotein granule / Signaling by RAF1 mutants / cell-cell junction / Signaling by moderate kinase activity BRAF mutants / Paradoxical activation of RAF signaling by kinase inactive BRAF / Signaling downstream of RAS mutants / Signaling by BRAF and RAF1 fusions / actin cytoskeleton / nucleosome Similarity search - Function | ||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) Amanita phalloides (death cap) Amanita phalloides (death cap) | ||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.65 Å | ||||||||||||
 Authors Authors | Boiero Sanders, M. / Oosterheert, W. / Hofnagel, O. / Bieling, P. / Raunser, S. | ||||||||||||
| Funding support |  Germany, European Union, 3items Germany, European Union, 3items
| ||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2024 Title: Phalloidin and DNase I-bound F-actin pointed end structures reveal principles of filament stabilization and disassembly. Authors: Micaela Boiero Sanders / Wout Oosterheert / Oliver Hofnagel / Peter Bieling / Stefan Raunser / Abstract: Actin filament turnover involves subunits binding to and dissociating from the filament ends, with the pointed end being the primary site of filament disassembly. Several molecules modulate filament ...Actin filament turnover involves subunits binding to and dissociating from the filament ends, with the pointed end being the primary site of filament disassembly. Several molecules modulate filament turnover, but the underlying mechanisms remain incompletely understood. Here, we present three cryo-EM structures of the F-actin pointed end in the presence and absence of phalloidin or DNase I. The two terminal subunits at the undecorated pointed end adopt a twisted conformation. Phalloidin can still bind and bridge these subunits, inducing a conformational shift to a flattened, F-actin-like state. This explains how phalloidin prevents depolymerization at the pointed end. Interestingly, two DNase I molecules simultaneously bind to the phalloidin-stabilized pointed end. In the absence of phalloidin, DNase I binding would disrupt the terminal actin subunit packing, resulting in filament disassembly. Our findings uncover molecular principles of pointed end regulation and provide structural insights into the kinetic asymmetry between the actin filament ends. | ||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9fjm.cif.gz | 265.3 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9fjm.ent.gz | 215.7 KB | Display | PDB format |
| PDBx/mmJSON format | 9fjm.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/fj/9fjmftp://data.pdbj.org/pub/pdb/validation_reports/fj/9fjm | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  50506MC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |



-Links
 PDBj
PDBj
















- Assembly
Assembly
| Deposited unit | 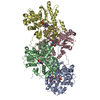
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 41632.422 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Details: Beta-actin recombinantly purified from insect cells. Source: (gene. exp.) Homo sapiens (human) / Gene: ACTB / Plasmid: p2336 pFL_ACTB_C272A / Cell line (production host): BTI-Tnao38 / Production host:  Trichoplusia ni (cabbage looper) / References: UniProt: P60709 Trichoplusia ni (cabbage looper) / References: UniProt: P60709#2: Protein/peptide | Type: Peptide-like / Class: Toxin / Mass: 808.899 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Details: phalloidin from Amanita phalloides. / Source: (natural) Amanita phalloides (death cap) / References: BIRD: PRD_002366#3: Chemical | ChemComp-ADP /   Mass: 427.201 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Feature type: SUBJECT OF INVESTIGATION / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Feature type: SUBJECT OF INVESTIGATION / Comment: ADP, energy-carrying molecule*YM#4: Chemical | ChemComp-MG /   Mass: 24.305 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION Mass: 24.305 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION#5: Chemical | ChemComp-PO4 / 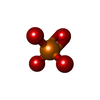  Mass: 94.971 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: PO4 / Feature type: SUBJECT OF INVESTIGATION Mass: 94.971 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: PO4 / Feature type: SUBJECT OF INVESTIGATIONHas ligand of interest | Y | |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight |
| ||||||||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||||||||
| Source (recombinant) | Organism: Trichoplusia ni (cabbage looper) / Cell: BTI-Tnao38 | ||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.1 Details: 1xKMEH (10 mM HEPES pH 7.1, 100 mM KCl, 2 mM MgCl2, 1 mM EGTA, 0.5 mM TCEP) | ||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||
| Specimen support | Grid material: GOLD / Grid mesh size: 200 divisions/in. / Grid type: Quantifoil R2/1 | ||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE-PROPANE / Humidity: 100 % / Chamber temperature: 286 K / Details: 3 seconds, force 0. |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS Details: 300 kV Titan Krios G2 microscope (Thermo Fisher Scientific) with an in-column Cs-corrector. |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 81000 X / Nominal defocus max: 2700 nm / Nominal defocus min: 1200 nm / Cs: 0.01 mm / C2 aperture diameter: 50 µm |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Electron dose: 64.6 e/Å2 / Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) / Num. of grids imaged: 1 / Num. of real images: 20393 |
| EM imaging optics | Energyfilter name: GIF Bioquantum / Details: Gatan energy filter. / Energyfilter slit width: 15 eV Spherical aberration corrector: Titan Krios G2 microscope (Thermo Fisher Scientific) with an in-column Cs-corrector. |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 5200600 / Details: Particles picked using SPHIRE-crYOLO. | ||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.65 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 280802 / Details: Local Refinement in CryoSPARC. / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL | ||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | PDB-ID: 8rtt Accession code: 8rtt / Details: actin and phalloidin model / Source name: PDB / Type: experimental model | ||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|

