 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8roh | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Human cohesin SMC3-HD(EQ)/RAD21-N complex - Apo conformation | ||||||||||||||||||||||||
 要素 要素 |
| ||||||||||||||||||||||||
 キーワード キーワード | NUCLEAR PROTEIN / 3D genome organization / Chromatin / Cohesin / ATPase activity / ATPase cycle | ||||||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報negative regulation of mitotic metaphase/anaphase transition / Cohesin Loading onto Chromatin / meiotic cohesin complex / Establishment of Sister Chromatid Cohesion / establishment of meiotic sister chromatid cohesion / cohesin complex / mitotic cohesin complex / positive regulation of sister chromatid cohesion / negative regulation of glial cell apoptotic process / negative regulation of G2/M transition of mitotic cell cycle ...negative regulation of mitotic metaphase/anaphase transition / Cohesin Loading onto Chromatin / meiotic cohesin complex / Establishment of Sister Chromatid Cohesion / establishment of meiotic sister chromatid cohesion / cohesin complex / mitotic cohesin complex / positive regulation of sister chromatid cohesion / negative regulation of glial cell apoptotic process / negative regulation of G2/M transition of mitotic cell cycle / mediator complex binding / replication-born double-strand break repair via sister chromatid exchange / establishment of mitotic sister chromatid cohesion / lateral element / chromatin looping / reciprocal meiotic recombination / microtubule motor activity / sister chromatid cohesion / negative regulation of interleukin-1 beta production / mitotic sister chromatid cohesion / stem cell population maintenance / mitotic spindle pole / lncRNA binding / dynein complex binding / regulation of DNA replication / positive regulation of interleukin-10 production / negative regulation of tumor necrosis factor production / chromosome, centromeric region / mitotic spindle assembly / SUMOylation of DNA damage response and repair proteins / beta-tubulin binding / cis-regulatory region sequence-specific DNA binding / protein localization to chromatin / Meiotic synapsis / Resolution of Sister Chromatid Cohesion / condensed nuclear chromosome / meiotic cell cycle / chromosome segregation / nuclear matrix / spindle pole / Separation of Sister Chromatids / double-strand break repair / mitotic cell cycle / chromosome / double-stranded DNA binding / midbody / DNA recombination / Estrogen-dependent gene expression / DNA-binding transcription factor binding / negative regulation of neuron apoptotic process / response to hypoxia / protein heterodimerization activity / cell division / DNA repair / apoptotic process / chromatin binding / regulation of transcription by RNA polymerase II / chromatin / ATP hydrolysis activity / nucleoplasm / ATP binding / nucleus / membrane / cytosol 類似検索 - 分子機能 | ||||||||||||||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||||||||||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 2.6 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 2.6 Å | ||||||||||||||||||||||||
 データ登録者 データ登録者 | Vitoria Gomes, M. / Romier, C. | ||||||||||||||||||||||||
| 資金援助 |  フランス, 7件 フランス, 7件
| ||||||||||||||||||||||||
 引用 引用 | ジャーナル: Cell Rep / 年: 2024 タイトル: The cohesin ATPase cycle is mediated by specific conformational dynamics and interface plasticity of SMC1A and SMC3 ATPase domains. 著者: Marina Vitoria Gomes / Pauline Landwerlin / Marie-Laure Diebold-Durand / Tajith B Shaik / Alexandre Durand / Edouard Troesch / Chantal Weber / Karl Brillet / Marianne Victoria Lemée / ...著者: Marina Vitoria Gomes / Pauline Landwerlin / Marie-Laure Diebold-Durand / Tajith B Shaik / Alexandre Durand / Edouard Troesch / Chantal Weber / Karl Brillet / Marianne Victoria Lemée / Christophe Decroos / Ludivine Dulac / Pierre Antony / Erwan Watrin / Eric Ennifar / Christelle Golzio / Christophe Romier / 要旨: Cohesin is key to eukaryotic genome organization and acts throughout the cell cycle in an ATP-dependent manner. The mechanisms underlying cohesin ATPase activity are poorly understood. Here, we ...Cohesin is key to eukaryotic genome organization and acts throughout the cell cycle in an ATP-dependent manner. The mechanisms underlying cohesin ATPase activity are poorly understood. Here, we characterize distinct steps of the human cohesin ATPase cycle and show that the SMC1A and SMC3 ATPase domains undergo specific but concerted structural rearrangements along this cycle. Specifically, whereas the proximal coiled coil of the SMC1A ATPase domain remains conformationally stable, that of the SMC3 displays an intrinsic flexibility. The ATP-dependent formation of the heterodimeric SMC1A/SMC3 ATPase module (engaged state) favors this flexibility, which is counteracted by NIPBL and DNA binding (clamped state). Opening of the SMC3/RAD21 interface (open-engaged state) stiffens the SMC3 proximal coiled coil, thus constricting together with that of SMC1A the ATPase module DNA-binding chamber. The plasticity of the ATP-dependent interface between the SMC1A and SMC3 ATPase domains enables these structural rearrangements while keeping the ATP gate shut. VIDEO ABSTRACT. | ||||||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8roh.cif.gz | 257.4 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8roh.ent.gz | 172.5 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8roh.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8roh_validation.pdf.gz | 459.3 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8roh_full_validation.pdf.gz | 465.6 KB | 表示 | |
| XML形式データ | 8roh_validation.xml.gz | 21.7 KB | 表示 | |
| CIF形式データ | 8roh_validation.cif.gz | 27.8 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/ro/8rohftp://data.pdbj.org/pub/pdb/validation_reports/ro/8roh | HTTPS FTP |
-関連構造データ
| 関連構造データ |  8p0aC  8pq5C  8ro6C  8ro7C  8ro8C  8ro9C  8roaC  8robC  8rocC  8rodC  8roeC  8rofC  8rogC  8roiC  8rojC  8rokC  8rolC C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |


-リンク
 PDBj
PDBj



- 集合体
集合体
| 登録構造単位 | 
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||
| 単位格子 |
|
-要素
| #1: タンパク質 | 分子量: 54448.754 Da / 分子数: 1 / 変異: E1144Q / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: SMC3, BAM, BMH, CSPG6, SMC3L1 / 発現宿主:  | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| #2: タンパク質 | 分子量: 12617.939 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RAD21, HR21, KIAA0078, NXP1, SCC1 / 発現宿主: | ||||||||
| #3: 化合物 | 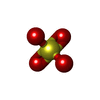  分子量: 96.063 Da / 分子数: 3 / 由来タイプ: 合成 / 式: SO4 分子量: 96.063 Da / 分子数: 3 / 由来タイプ: 合成 / 式: SO4#4: 化合物 | ChemComp-GOL / | 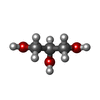  分子量: 92.094 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C3H8O3 分子量: 92.094 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C3H8O3#5: 水 | ChemComp-HOH / |  分子量: 18.015 Da / 分子数: 9 / 由来タイプ: 天然 / 式: H2O 分子量: 18.015 Da / 分子数: 9 / 由来タイプ: 天然 / 式: H2O研究の焦点であるリガンドがあるか | N | Has protein modification | N | |
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 3.64 Å3/Da / 溶媒含有率: 66.2 % |
|---|---|
| 結晶化 | 温度: 293 K / 手法: 蒸気拡散法, シッティングドロップ法 詳細: 0.21M NaSO4 ; 0.1M Bis-tris propane pH 7; 16% w/v PEG3350 |
-データ収集
| 回折 | 平均測定温度: 100 K / Serial crystal experiment: N |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: SLS  / ビームライン: X06DA / 波長: 1.000031 Å / ビームライン: X06DA / 波長: 1.000031 Å |
| 検出器 | タイプ: DECTRIS PILATUS 2M-F / 検出器: PIXEL / 日付: 2020年8月28日 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 1.000031 Å / 相対比: 1 |
| 反射 | 解像度: 2.6→50 Å / Num. obs: 31414 / % possible obs: 99.9 % / 冗長度: 26.2 % / Biso Wilson estimate: 60.16 Å2 / CC1/2: 0.999 / Rmerge(I) obs: 0.28 / Net I/σ(I): 14.93 |
| 反射 シェル | 解像度: 2.6→2.76 Å / 冗長度: 25.7 % / Rmerge(I) obs: 3.842 / Mean I/σ(I) obs: 1.02 / Num. unique obs: 4948 / CC1/2: 0.366 / % possible all: 99.7 |
- 解析
解析
| ソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 / 解像度: 2.6→49.6 Å / SU ML: 0.4392 / 交差検証法: FREE R-VALUE / σ(F): 0.48 / 位相誤差: 30.9996 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å / 溶媒モデル: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 71.92 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 2.6→49.6 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ | Refine-ID: X-RAY DIFFRACTION
|
