 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8rok: Human cohesin SMC3-HD(EQ)/RAD21-N complex - ATP-Mg-bound conforma... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8rok | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Human cohesin SMC3-HD(EQ)/RAD21-N complex - ATP-Mg-bound conformation - Form 1 | ||||||||||||||||||||||||
 Components Components |
| ||||||||||||||||||||||||
 Keywords Keywords | NUCLEAR PROTEIN / 3D genome organization / Chromatin / Cohesin / ATPase activity / ATPase cycle | ||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationmitotic cohesin complex / negative regulation of mitotic metaphase/anaphase transition / meiotic cohesin complex / Cohesin Loading onto Chromatin / Establishment of Sister Chromatid Cohesion / cohesin complex / establishment of meiotic sister chromatid cohesion / positive regulation of sister chromatid cohesion / replication-born double-strand break repair via sister chromatid exchange / negative regulation of glial cell apoptotic process ...mitotic cohesin complex / negative regulation of mitotic metaphase/anaphase transition / meiotic cohesin complex / Cohesin Loading onto Chromatin / Establishment of Sister Chromatid Cohesion / cohesin complex / establishment of meiotic sister chromatid cohesion / positive regulation of sister chromatid cohesion / replication-born double-strand break repair via sister chromatid exchange / negative regulation of glial cell apoptotic process / negative regulation of G2/M transition of mitotic cell cycle / mediator complex binding / establishment of mitotic sister chromatid cohesion / lateral element / reciprocal meiotic recombination / mitotic sister chromatid cohesion / microtubule motor activity / sister chromatid cohesion / negative regulation of interleukin-1 beta production / mitotic spindle pole / lncRNA binding / regulation of DNA replication / positive regulation of interleukin-10 production / dynein complex binding / mitotic spindle assembly / negative regulation of tumor necrosis factor production / chromosome, centromeric region / SUMOylation of DNA damage response and repair proteins / cis-regulatory region sequence-specific DNA binding / beta-tubulin binding / protein localization to chromatin / Resolution of Sister Chromatid Cohesion / Meiotic synapsis / condensed nuclear chromosome / chromosome segregation / meiotic cell cycle / nuclear matrix / spindle pole / Separation of Sister Chromatids / double-strand break repair / mitotic cell cycle / chromosome / double-stranded DNA binding / DNA recombination / negative regulation of neuron apoptotic process / Estrogen-dependent gene expression / DNA-binding transcription factor binding / response to hypoxia / protein heterodimerization activity / cell division / DNA repair / chromatin binding / apoptotic process / regulation of transcription by RNA polymerase II / chromatin / ATP hydrolysis activity / DNA-templated transcription / nucleoplasm / ATP binding / membrane / nucleus / cytosol Similarity search - Function | ||||||||||||||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||||||||||||||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.25 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.25 Å | ||||||||||||||||||||||||
 Authors Authors | Vitoria Gomes, M. / Romier, C. | ||||||||||||||||||||||||
| Funding support |  France, 7items France, 7items
| ||||||||||||||||||||||||
 Citation Citation | Journal: Cell Rep / Year: 2024 Title: The cohesin ATPase cycle is mediated by specific conformational dynamics and interface plasticity of SMC1A and SMC3 ATPase domains. Authors: Marina Vitoria Gomes / Pauline Landwerlin / Marie-Laure Diebold-Durand / Tajith B Shaik / Alexandre Durand / Edouard Troesch / Chantal Weber / Karl Brillet / Marianne Victoria Lemée / ...Authors: Marina Vitoria Gomes / Pauline Landwerlin / Marie-Laure Diebold-Durand / Tajith B Shaik / Alexandre Durand / Edouard Troesch / Chantal Weber / Karl Brillet / Marianne Victoria Lemée / Christophe Decroos / Ludivine Dulac / Pierre Antony / Erwan Watrin / Eric Ennifar / Christelle Golzio / Christophe Romier / Abstract: Cohesin is key to eukaryotic genome organization and acts throughout the cell cycle in an ATP-dependent manner. The mechanisms underlying cohesin ATPase activity are poorly understood. Here, we ...Cohesin is key to eukaryotic genome organization and acts throughout the cell cycle in an ATP-dependent manner. The mechanisms underlying cohesin ATPase activity are poorly understood. Here, we characterize distinct steps of the human cohesin ATPase cycle and show that the SMC1A and SMC3 ATPase domains undergo specific but concerted structural rearrangements along this cycle. Specifically, whereas the proximal coiled coil of the SMC1A ATPase domain remains conformationally stable, that of the SMC3 displays an intrinsic flexibility. The ATP-dependent formation of the heterodimeric SMC1A/SMC3 ATPase module (engaged state) favors this flexibility, which is counteracted by NIPBL and DNA binding (clamped state). Opening of the SMC3/RAD21 interface (open-engaged state) stiffens the SMC3 proximal coiled coil, thus constricting together with that of SMC1A the ATPase module DNA-binding chamber. The plasticity of the ATP-dependent interface between the SMC1A and SMC3 ATPase domains enables these structural rearrangements while keeping the ATP gate shut. VIDEO ABSTRACT. | ||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8rok.cif.gz | 474.1 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8rok.ent.gz | 322.5 KB | Display | PDB format |
| PDBx/mmJSON format | 8rok.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ro/8rokftp://data.pdbj.org/pub/pdb/validation_reports/ro/8rok | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  8p0aC  8pq5C  8ro6C  8ro7C  8ro8C  8ro9C  8roaC  8robC  8rocC  8rodC  8roeC  8rofC  8rogC  8rohC  8roiC  8rojC  8rolC C: citing same article ( |
|---|---|
| Similar structure data |


-Links
 PDBj
PDBj




- Assembly
Assembly
| Deposited unit | 
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||||||
| 2 | 
| ||||||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 52391.367 Da / Num. of mol.: 2 / Mutation: E1144Q Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: SMC3, BAM, BMH, CSPG6, SMC3L1 / Production host:  #2: Protein | Mass: 12617.939 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: RAD21, HR21, KIAA0078, NXP1, SCC1 / Production host: #3: Chemical | 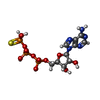  Mass: 523.247 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O12P3S / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP-gamma-S, energy-carrying molecule analogue*YM Mass: 523.247 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O12P3S / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP-gamma-S, energy-carrying molecule analogue*YM#4: Chemical |   Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION#5: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 90 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 90 / Source method: isolated from a natural source / Formula: H2OHas ligand of interest | Y | Has protein modification | N | |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.42 Å3/Da / Density % sol: 49.15 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, sitting drop Details: 0.3 M NaCl; 0.05 M L-Arginine; 0.1 M Tris pH7.5; 22.5 % v/v PEG Smear Broad; 0.05 M L-Glutamic acid monosodium salt hydrate |
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: SLS  / Beamline: X06DA / Wavelength: 1.000002 Å / Beamline: X06DA / Wavelength: 1.000002 Å |
| Detector | Type: DECTRIS PILATUS 2M-F / Detector: PIXEL / Date: Nov 22, 2019 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1.000002 Å / Relative weight: 1 |
| Reflection | Resolution: 2.25→50 Å / Num. obs: 58135 / % possible obs: 98 % / Redundancy: 13.6 % / Biso Wilson estimate: 48.58 Å2 / CC1/2: 1 / Rmerge(I) obs: 0.098 / Net I/σ(I): 20.32 |
| Reflection shell | Resolution: 2.25→2.39 Å / Redundancy: 13.1 % / Rmerge(I) obs: 2.233 / Mean I/σ(I) obs: 1.34 / Num. unique obs: 8944 / CC1/2: 0.816 / % possible all: 94.2 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT / Resolution: 2.25→47.21 Å / SU ML: 0.3738 / Cross valid method: FREE R-VALUE / σ(F): 0.38 / Phase error: 33.5302 Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 74.7 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.25→47.21 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|
