 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8eza | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | NHEJ Long-range complex with PAXX | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | DNA BINDING PROTEIN/DNA / Complex / Kinase / NHEJ / DNA BINDING PROTEIN / DNA BINDING PROTEIN-DNA complex | |||||||||
| Function / homology |  Function and homology information Function and homology informationT cell receptor V(D)J recombination / FHA domain binding / positive regulation of chromosome organization / pro-B cell differentiation / DNA ligase IV complex / positive regulation of ligase activity / positive regulation of platelet formation / DNA ligase activity / DNA-dependent protein kinase complex / DNA double-strand break attachment to nuclear envelope ...T cell receptor V(D)J recombination / FHA domain binding / positive regulation of chromosome organization / pro-B cell differentiation / DNA ligase IV complex / positive regulation of ligase activity / positive regulation of platelet formation / DNA ligase activity / DNA-dependent protein kinase complex / DNA double-strand break attachment to nuclear envelope / Ku70:Ku80 complex / DNA ligase (ATP) / negative regulation of t-circle formation / DNA-dependent protein kinase activity / DNA end binding / DNA ligase (ATP) activity / small-subunit processome assembly / positive regulation of lymphocyte differentiation / histone H2AXS139 kinase activity / DNA-dependent protein kinase-DNA ligase 4 complex / immunoglobulin V(D)J recombination / V(D)J recombination / nonhomologous end joining complex / nucleotide-excision repair, DNA gap filling / cellular response to X-ray / isotype switching / cellular response to lithium ion / regulation of epithelial cell proliferation / regulation of smooth muscle cell proliferation / double-strand break repair via classical nonhomologous end joining / double-strand break repair via alternative nonhomologous end joining / Cytosolic sensors of pathogen-associated DNA / protein localization to site of double-strand break / nuclear telomere cap complex / single strand break repair / telomere capping / IRF3-mediated induction of type I IFN / regulation of hematopoietic stem cell differentiation / positive regulation of neurogenesis / U3 snoRNA binding / regulation of telomere maintenance / recombinational repair / cellular hyperosmotic salinity response / somatic stem cell population maintenance / protein localization to chromosome, telomeric region / maturation of 5.8S rRNA / positive regulation of double-strand break repair via nonhomologous end joining / DNA biosynthetic process / 2-LTR circle formation / negative regulation of cGAS/STING signaling pathway / response to ionizing radiation / peptidyl-threonine phosphorylation / telomeric repeat DNA binding / ligase activity / negative regulation of protein phosphorylation / T cell differentiation / DNA 3'-5' helicase / 5'-deoxyribose-5-phosphate lyase activity / chromosome organization / response to X-ray / 3'-5' DNA helicase activity / ATP-dependent activity, acting on DNA / telomere maintenance via telomerase / SUMOylation of DNA damage response and repair proteins / condensed chromosome / mitotic G1 DNA damage checkpoint signaling / site of DNA damage / B cell differentiation / DNA polymerase binding / positive regulation of erythrocyte differentiation / response to gamma radiation / activation of innate immune response / telomere maintenance / cyclin binding / DNA helicase activity / protein modification process / DNA-(apurinic or apyrimidinic site) lyase / class I DNA-(apurinic or apyrimidinic site) endonuclease activity / cellular response to ionizing radiation / positive regulation of translation / central nervous system development / peptidyl-serine phosphorylation / small-subunit processome / Nonhomologous End-Joining (NHEJ) / cellular response to gamma radiation / cell population proliferation / protein-DNA complex / base-excision repair / establishment of integrated proviral latency / intrinsic apoptotic signaling pathway in response to DNA damage / regulation of circadian rhythm / double-strand break repair via nonhomologous end joining / positive regulation of fibroblast proliferation / cellular response to insulin stimulus / enzyme activator activity / in utero embryonic development / fibrillar center / T cell differentiation in thymus / neuron apoptotic process / rhythmic process Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4.39 Å | |||||||||
 Authors Authors | Chen, S. / He, Y. | |||||||||
| Funding support |  United States, 2items United States, 2items
| |||||||||
 Citation Citation | Journal: Sci Adv / Year: 2023 Title: Cryo-EM visualization of DNA-PKcs structural intermediates in NHEJ. Authors: Siyu Chen / Alex Vogt / Linda Lee / Tasmin Naila / Ryan McKeown / Alan E Tomkinson / Susan P Lees-Miller / Yuan He /  Abstract: DNA double-strand breaks (DSBs), one of the most cytotoxic forms of DNA damage, can be repaired by the tightly regulated nonhomologous end joining (NHEJ) machinery (Stinson and Loparo and Zhao ). ...DNA double-strand breaks (DSBs), one of the most cytotoxic forms of DNA damage, can be repaired by the tightly regulated nonhomologous end joining (NHEJ) machinery (Stinson and Loparo and Zhao ). Core NHEJ factors form an initial long-range (LR) synaptic complex that transitions into a DNA-PKcs (DNA-dependent protein kinase, catalytic subunit)-free, short-range state to align the DSB ends (Chen ). Using single-particle cryo-electron microscopy, we have visualized three additional key NHEJ complexes representing different transition states, with DNA-PKcs adopting distinct dimeric conformations within each of them. Upon DNA-PKcs autophosphorylation, the LR complex undergoes a substantial conformational change, with both Ku and DNA-PKcs rotating outward to promote DNA break exposure and DNA-PKcs dissociation. We also captured a dimeric state of catalytically inactive DNA-PKcs, which resembles structures of other PIKK (Phosphatidylinositol 3-kinase-related kinase) family kinases, revealing a model of the full regulatory cycle of DNA-PKcs during NHEJ. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8eza.cif.gz | 2.1 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8eza.ent.gz | 1.7 MB | Display | PDB format |
| PDBx/mmJSON format | 8eza.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ez/8ezaftp://data.pdbj.org/pub/pdb/validation_reports/ez/8eza | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  28732MC  8ez9C 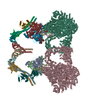 8ezbC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |







-Links
 PDBj
PDBj












































- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 5 types, 12 molecules STLCHIFGOPXY
| #1: Protein | Mass: 21663.498 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PAXX, C9orf142, XLS / Production host:  #4: Protein | Mass: 469673.219 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: HELA / References: UniProt: P78527#8: Protein | Mass: 33372.234 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: NHEJ1, XLF / Production host: #9: Protein | Mass: 38337.703 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: XRCC4 / Production host: #10: Protein | Mass: 104124.953 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: LIG4 / Production host: |
|---|
-X-ray repair cross-complementing protein ... , 2 types, 4 molecules JAKB
| #2: Protein | Mass: 69945.039 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Production host:  unidentified baculovirus / References: UniProt: P12956 unidentified baculovirus / References: UniProt: P12956#3: Protein | Mass: 82812.438 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Production host: unidentified baculovirus / References: UniProt: P13010 |
|---|
-DNA chain , 2 types, 4 molecules MDNE
| #6: DNA chain | Mass: 9510.159 Da / Num. of mol.: 2 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human)#7: DNA chain | Mass: 9236.976 Da / Num. of mol.: 2 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) |
|---|
-Protein/peptide / Non-polymers , 2 types, 4 molecules RQ
| #11: Chemical |  Mass: 507.181 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP, energy-carrying molecule*YM#5: Protein/peptide | Mass: 1720.111 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: HELA / References: non-specific serine/threonine protein kinase |
|---|
-Details
| Has ligand of interest | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: NHEJ Long-range complex with PAXX / Type: COMPLEX / Entity ID: #1-#7, #9-#10, #8 / Source: MULTIPLE SOURCES |
|---|---|
| Molecular weight | Value: 1.75 MDa / Experimental value: NO |
| Buffer solution | pH: 7.9 |
| Specimen | Conc.: 2 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Grid material: COPPER / Grid mesh size: 200 divisions/in. / Grid type: Quantifoil R2/1 |
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 302 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 60000 X / Calibrated magnification: 60000 X / Nominal defocus max: 4000 nm / Nominal defocus min: 2000 nm / Calibrated defocus min: 2000 nm / Calibrated defocus max: 4000 nm / Cs: 2 mm / C2 aperture diameter: 100 µm / Alignment procedure: BASIC |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER / Temperature (max): 70 K / Temperature (min): 70 K |
| Image recording | Average exposure time: 4 sec. / Electron dose: 65 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) / Num. of grids imaged: 1 / Num. of real images: 11006 |
| Image scans | Width: 5760 / Height: 4092 |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 1101255 | ||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C2 (2 fold cyclic) | ||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4.39 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 138252 / Algorithm: FOURIER SPACE / Num. of class averages: 1 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL / Target criteria: Cross-correlation coefficient |
