National Institutes of Health/Eunice Kennedy Shriver National Institute of Child Health & Human Development (NIH/NICHD)
R01GM125885
米国
National Institutes of Health/Eunice Kennedy Shriver National Institute of Child Health & Human Development (NIH/NICHD)
P30CA021765
米国
引用
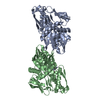
ジャーナル: Mol Cell / 年: 2023 タイトル: E3 ligase autoinhibition by C-degron mimicry maintains C-degron substrate fidelity. 著者: Daniel C Scott / Moeko T King / Kheewoong Baek / Clifford T Gee / Ravi Kalathur / Jerry Li / Nicholas Purser / Amanda Nourse / Sergio C Chai / Sivaraja Vaithiyalingam / Taosheng Chen / ...著者: Daniel C Scott / Moeko T King / Kheewoong Baek / Clifford T Gee / Ravi Kalathur / Jerry Li / Nicholas Purser / Amanda Nourse / Sergio C Chai / Sivaraja Vaithiyalingam / Taosheng Chen / Richard E Lee / Stephen J Elledge / Gary Kleiger / Brenda A Schulman / 要旨: E3 ligase recruitment of proteins containing terminal destabilizing motifs (degrons) is emerging as a major form of regulation. How those E3s discriminate bona fide substrates from other proteins ...E3 ligase recruitment of proteins containing terminal destabilizing motifs (degrons) is emerging as a major form of regulation. How those E3s discriminate bona fide substrates from other proteins with terminal degron-like sequences remains unclear. Here, we report that human KLHDC2, a CRL2 substrate receptor targeting C-terminal Gly-Gly degrons, is regulated through interconversion between two assemblies. In the self-inactivated homotetramer, KLHDC2's C-terminal Gly-Ser motif mimics a degron and engages the substrate-binding domain of another protomer. True substrates capture the monomeric CRL2, driving E3 activation by neddylation and subsequent substrate ubiquitylation. Non-substrates such as NEDD8 bind KLHDC2 with high affinity, but its slow on rate prevents productive association with CRL2. Without substrate, neddylated CRL2 assemblies are deactivated via distinct mechanisms: the monomer by deneddylation and the tetramer by auto-ubiquitylation. Thus, substrate specificity is amplified by KLHDC2 self-assembly acting like a molecular timer, where only bona fide substrates may bind before E3 ligase inactivation.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) X線回折 /
X線回折 /  データ登録者
データ登録者 米国, 2件
米国, 2件  引用
引用
 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード


 PDBj
PDBj 集合体
集合体


 分子量: 18.015 Da / 分子数: 403 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 403 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 解析
解析