 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8bqu | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
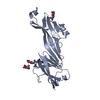
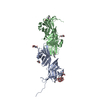
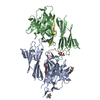
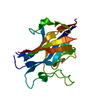
| タイトル | Molecular basis of ZP3/ZP1 heteropolymerization: crystal structure of a native vertebrate egg coat filament | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | STRUCTURAL PROTEIN / Cell adhesion / fertilization / egg-sperm interaction / gamete recognition / sperm receptor / extracellular matrix / egg coat / zona pellucida / vitelline envelope / fish chorion / glycoprotein / N-glycan / ZP module / ZP-N domain / ZP_C domain / trefoil domain / medaka / Japanese rice fish | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報egg coat formation / structural constituent of egg coat / egg coat / positive regulation of acrosome reaction / binding of sperm to zona pellucida / single fertilization / plasma membrane 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  | ||||||||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 2.7 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 2.7 Å | ||||||||||||
 データ登録者 データ登録者 | Bokhove, M. / de Sanctis, D. / Yasumasu, S. / Jovine, L. | ||||||||||||
| 資金援助 |  スウェーデン, 3件 スウェーデン, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Cell / 年: 2024 タイトル: ZP2 cleavage blocks polyspermy by modulating the architecture of the egg coat. 著者: Shunsuke Nishio / Chihiro Emori / Benjamin Wiseman / Dirk Fahrenkamp / Elisa Dioguardi / Sara Zamora-Caballero / Marcel Bokhove / Ling Han / Alena Stsiapanava / Blanca Algarra / Yonggang Lu / ...著者: Shunsuke Nishio / Chihiro Emori / Benjamin Wiseman / Dirk Fahrenkamp / Elisa Dioguardi / Sara Zamora-Caballero / Marcel Bokhove / Ling Han / Alena Stsiapanava / Blanca Algarra / Yonggang Lu / Mayo Kodani / Rachel E Bainbridge / Kayla M Komondor / Anne E Carlson / Michael Landreh / Daniele de Sanctis / Shigeki Yasumasu / Masahito Ikawa / Luca Jovine /    要旨: Following the fertilization of an egg by a single sperm, the egg coat or zona pellucida (ZP) hardens and polyspermy is irreversibly blocked. These events are associated with the cleavage of the N- ...Following the fertilization of an egg by a single sperm, the egg coat or zona pellucida (ZP) hardens and polyspermy is irreversibly blocked. These events are associated with the cleavage of the N-terminal region (NTR) of glycoprotein ZP2, a major subunit of ZP filaments. ZP2 processing is thought to inactivate sperm binding to the ZP, but its molecular consequences and connection with ZP hardening are unknown. Biochemical and structural studies show that cleavage of ZP2 triggers its oligomerization. Moreover, the structure of a native vertebrate egg coat filament, combined with AlphaFold predictions of human ZP polymers, reveals that two protofilaments consisting of type I (ZP3) and type II (ZP1/ZP2/ZP4) components interlock into a left-handed double helix from which the NTRs of type II subunits protrude. Together, these data suggest that oligomerization of cleaved ZP2 NTRs extensively cross-links ZP filaments, rigidifying the egg coat and making it physically impenetrable to sperm. #1: ジャーナル: J Mol Biol / 年: 1985 タイトル: Mouse egg extracellular coat is a matrix of interconnected filaments possessing a structural repeat. 著者: Greve, J.M. / Wassarman, P.M. #2: ジャーナル: J Biochem / 年: 1989 タイトル: Purification and partial characterization of high choriolytic enzyme (HCE), a component of the hatching enzyme of the teleost, Oryzias latipes. 著者: Yasumasu, S. / Yamagami, K. / Iuchi, I. #3: ジャーナル: J Biochem / 年: 1989 タイトル: Isolation and some properties of low choriolytic enzyme (LCE), a component of the hatching enzyme of the teleost, Oryzias latipes. 著者: Yasumasu, S. / Iuchi, I. / Yamagami, K. #4: ジャーナル: Dev Biol / 年: 1992 タイトル: Isolation of cDNAs for LCE and HCE, two constituent proteases of the hatching enzyme of Oryzias latipes, and concurrent expression of their mRNAs during development. 著者: Yasumasu, S. / Yamada, K. / Akasaka, K. / Mitsunaga, K. / Iuchi, I. / Shimada, H. / Yamagami, K. #5: ジャーナル: Proc Natl Acad Sci U S A / 年: 1997 タイトル: Cloning of cDNA and estrogen-induced hepatic gene expression for choriogenin H, a precursor protein of the fish egg envelope (chorion). 著者: Murata, K. / Sugiyama, H. / Yasumasu, S. / Iuchi, I. / Yasumasu, I. / Yamagami, K. #6: ジャーナル: Dev Biol / 年: 1995 タイトル: Cloning of cDNAs for the precursor protein of a low-molecular-weight subunit of the inner layer of the egg envelope (chorion) of the fish Oryzias latipes. 著者: Murata, K. / Sasaki, T. / Yasumasu, S. / Iuchi, I. / Enami, J. / Yasumasu, I. / Yamagami, K. #7: ジャーナル: Dev Growth Differ / 年: 1998 タイトル: The third egg envelope subunit in fish: cDNA cloning and analysis, and gene expression. 著者: Sugiyama, H. / Yasumasu, S. / Murata, K. / Iuchi, I. / Yamagami, K. #8: ジャーナル: J Biochem / 年: 1999 タイトル: Formation of mature egg envelope subunit proteins from their precursors (choriogenins) in the fish, Oryzias latipes: loss of partial C-terminal sequences of the choriogenins. 著者: Sugiyama, H. / Murata, K. / Iuchi, I. / Nomura, K. / Yamagami, K. #9: ジャーナル: Nature / 年: 2008タイトル: Crystal structure of the ZP-N domain of ZP3 reveals the core fold of animal egg coats. 著者: Monne, M. / Han, L. / Schwend, T. / Burendahl, S. / Jovine, L. #10: ジャーナル: J Biochem / 年: 2010 タイトル: Mechanism of egg envelope digestion by hatching enzymes, HCE and LCE in medaka, Oryzias latipes. 著者: Yasumasu, S. / Kawaguchi, M. / Ouchi, S. / Sano, K. / Murata, K. / Sugiyama, H. / Akema, T. / Iuchi, I. #11: ジャーナル: Cell / 年: 2010タイトル: Insights into egg coat assembly and egg-sperm interaction from the X-ray structure of full-length ZP3. 著者: Ling Han / Magnus Monné / Hiroki Okumura / Thomas Schwend / Amy L Cherry / David Flot / Tsukasa Matsuda / Luca Jovine / 要旨: ZP3, a major component of the zona pellucida (ZP) matrix coating mammalian eggs, is essential for fertilization by acting as sperm receptor. By retaining a propeptide that contains a polymerization- ...ZP3, a major component of the zona pellucida (ZP) matrix coating mammalian eggs, is essential for fertilization by acting as sperm receptor. By retaining a propeptide that contains a polymerization-blocking external hydrophobic patch (EHP), we determined the crystal structure of an avian homolog of ZP3 at 2.0 Å resolution. The structure unveils the fold of a complete ZP domain module in a homodimeric arrangement required for secretion and reveals how EHP prevents premature incorporation of ZP3 into the ZP. This suggests mechanisms underlying polymerization and how local structural differences, reflected by alternative disulfide patterns, control the specificity of ZP subunit interaction. Close relative positioning of a conserved O-glycan important for sperm binding and the hypervariable, positively selected C-terminal region of ZP3 suggests a concerted role in the regulation of species-restricted gamete recognition. Alternative conformations of the area around the O-glycan indicate how sperm binding could trigger downstream events via intramolecular signaling. #12: ジャーナル: Curr Top Dev Biol / 年: 2018 タイトル: Structure of Zona Pellucida Module Proteins. 著者: Marcel Bokhove / Luca Jovine / 要旨: The egg coat, an extracellular matrix made up of glycoprotein filaments, plays a key role in animal fertilization by acting as a gatekeeper for sperm. Egg coat components polymerize using a common ...The egg coat, an extracellular matrix made up of glycoprotein filaments, plays a key role in animal fertilization by acting as a gatekeeper for sperm. Egg coat components polymerize using a common zona pellucida (ZP) "domain" module that consists of two related immunoglobulin-like domains, called ZP-N and ZP-C. The ZP module has also been recognized in a large number of other secreted proteins with different biological functions, whose mutations are linked to severe human diseases. During the last decade, tremendous progress has been made toward understanding the atomic architecture of the ZP module and the structural basis of its polymerization. Moreover, sperm-binding regions at the N-terminus of mollusk and mammalian egg coat subunits were found to consist of domain repeats that also adopt a ZP-N fold. This discovery revealed an unexpected link between invertebrate and vertebrate fertilization and led to the first structure of an egg coat-sperm protein recognition complex. In this review we summarize these exciting findings, discuss their functional implications, and outline future challenges that must be addressed in order to develop a comprehensive view of this family of biomedically important extracellular molecules. #13: ジャーナル: EMBO J / 年: 2020タイトル: Cryo-EM structure of native human uromodulin, a zona pellucida module polymer. 著者: Alena Stsiapanava / Chenrui Xu / Martina Brunati / Sara Zamora-Caballero / Céline Schaeffer / Marcel Bokhove / Ling Han / Hans Hebert / Marta Carroni / Shigeki Yasumasu / Luca Rampoldi / Bin ...著者: Alena Stsiapanava / Chenrui Xu / Martina Brunati / Sara Zamora-Caballero / Céline Schaeffer / Marcel Bokhove / Ling Han / Hans Hebert / Marta Carroni / Shigeki Yasumasu / Luca Rampoldi / Bin Wu / Luca Jovine /   要旨: Assembly of extracellular filaments and matrices mediating fundamental biological processes such as morphogenesis, hearing, fertilization, and antibacterial defense is driven by a ubiquitous ...Assembly of extracellular filaments and matrices mediating fundamental biological processes such as morphogenesis, hearing, fertilization, and antibacterial defense is driven by a ubiquitous polymerization module known as zona pellucida (ZP) "domain". Despite the conservation of this element from hydra to humans, no detailed information is available on the filamentous conformation of any ZP module protein. Here, we report a cryo-electron microscopy study of uromodulin (UMOD)/Tamm-Horsfall protein, the most abundant protein in human urine and an archetypal ZP module-containing molecule, in its mature homopolymeric state. UMOD forms a one-start helix with an unprecedented 180-degree twist between subunits enfolded by interdomain linkers that have completely reorganized as a result of propeptide dissociation. Lateral interaction between filaments in the urine generates sheets exposing a checkerboard of binding sites to capture uropathogenic bacteria, and UMOD-based models of heteromeric vertebrate egg coat filaments identify a common sperm-binding region at the interface between subunits. #14: ジャーナル: Zoological Lett / 年: 2022 タイトル: Targeted deletion of liver-expressed Choriogenin L results in the production of soft eggs and infertility in medaka, Oryzias latipes. 著者: Murata, K. / Kinoshita, M. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8bqu.cif.gz | 420.2 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8bqu.ent.gz | 279.1 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8bqu.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/bq/8bquftp://data.pdbj.org/pub/pdb/validation_reports/bq/8bqu | HTTPS FTP |
|---|
-関連構造データ







-リンク
 PDBj
PDBj- 集合体
集合体
| 登録構造単位 | 
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||
| 単位格子 |
|
-要素
| #1: タンパク質 | 分子量: 34873.172 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: ZP-N domain + interdomain linker + ZP-C domain / 由来: (天然) |
|---|---|
| #2: タンパク質 | 分子量: 18734.260 Da / 分子数: 1 / 由来タイプ: 天然 詳細: C-terminal half of the ZP-N/ZP-C interdomain linker + ZP-C domain 由来: (天然) |
| #3: タンパク質 | 分子量: 17876.166 Da / 分子数: 1 / 由来タイプ: 天然 詳細: Trefoil domain + ZP-N domain + N-terminal half of the ZP-N/ZP-C interdomain linker 由来: (天然) |
| #4: 多糖 | beta-D-galactopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-2)-alpha-D-mannopyranose- ...beta-D-galactopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-2)-alpha-D-mannopyranose-(1-3)-[alpha-D-mannopyranose-(1-6)]beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose タイプ: oligosaccharide / 分子量: 1276.157 Da / 分子数: 1 / 由来タイプ: 組換発現 |
| 研究の焦点であるリガンドがあるか | Y |
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 3.46 Å3/Da / 溶媒含有率: 64.16 % / 解説: Bipyramidal prismatic |
|---|---|
| 結晶化 | 温度: 277.15 K / 手法: 蒸気拡散法 / pH: 8 詳細: 22% (w/v) PEG 3350, 0.2 M sodium/potassium phosphate, 0.1 M Tris-HCl pH 8.0 |
-データ収集
| 回折 | 平均測定温度: 100 K / Serial crystal experiment: N | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: Diamond  / ビームライン: I02 / 波長: 0.97938 Å / ビームライン: I02 / 波長: 0.97938 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 検出器 | タイプ: DECTRIS PILATUS 6M-F / 検出器: PIXEL / 日付: 2014年11月29日 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 放射波長 | 波長: 0.97938 Å / 相対比: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 反射 | 解像度: 2.419→51.147 Å / Num. obs: 14348 / % possible obs: 89.9 % / 冗長度: 4.39 % 詳細: Some remarks regarding the mmCIF items written, the PDB Exchange Dictionary (PDBx/mmCIF) Version 5.0 supporting the data files in the current PDB archive (dictionary version 5.325, last ...詳細: Some remarks regarding the mmCIF items written, the PDB Exchange Dictionary (PDBx/mmCIF) Version 5.0 supporting the data files in the current PDB archive (dictionary version 5.325, last updated 2020-04-13: http://mmcif.wwpdb.org/dictionaries/mmcif_pdbx_v50.dic/Index/) and the actual quantities provided by MRFANA (https://github.com/githubgphl/MRFANA) from the autoPROC package (https://www.globalphasing.com/autoproc/). In general, the mmCIF categories here should provide items that are currently used in the PDB archive. If there are alternatives, the one recommended by the PDB developers has been selected. The distinction between *_all and *_obs quantities is not always clear: often only one version is actively used within the PDB archive (or is the one recommended by PDB developers). The intention of distinguishing between classes of reflections before and after some kind of observation criterion was applied, can in principle be useful - but such criteria change in various ways throughout the data processing steps (rejection of overloaded or too partial reflections, outlier/misfit rejections during scaling etc) and there is no retrospect computation of data scaling/merging statistics for the reflections used in the final refinement (where another observation criterion might have been applied). Typical data processing will usually only provide one version of statistics at various stages and these are given in the recommended item here, irrespective of the "_all" and "_obs" connotation, see e.g. the use of _reflns.pdbx_Rmerge_I_obs, _reflns.pdbx_Rrim_I_all and _reflns.pdbx_Rpim_I_all. Please note that all statistics related to "merged intensities" (or "merging") are based on inverse-variance weighting of the individual measurements making up a symmetry-unique reflection. This is standard for several decades now, even if some of the dictionary definitions seem to suggest that a simple "mean" or "average" intensity is being used instead. R-values are always given for all symmetry-equivalent reflections following Friedel's law, i.e. Bijvoet pairs are not treated separately (since we want to describe the overall mean intensity and not the mean I(+) and I(-) here). The Rrim metric is identical to the Rmeas R-value and only differs in name. _reflns.pdbx_number_measured_all is the number of measured intensities just before the final merging step (at which point no additional rejection takes place). _reflns.number_obs is the number of symmetry-unique observations, i.e. the result of merging those measurements via inverse-variance weighting. _reflns.pdbx_netI_over_sigmaI is based on the merged intensities (_reflns.number_obs) as expected. _reflns.pdbx_redundancy is synonymous with "multiplicity". The per-shell item _reflns_shell.number_measured_all corresponds to the overall value _reflns.pdbx_number_measured_all. The per-shell item _reflns_shell.number_unique_all corresponds to the overall value _reflns.number_obs. The per-shell item _reflns_shell.percent_possible_all corresponds to the overall value _reflns.percent_possible_obs. The per-shell item _reflns_shell.meanI_over_sigI_obs corresponds to the overall value given as _reflns.pdbx_netI_over_sigmaI. But be aware of the incorrect definition of the former in the current dictionary! CC1/2: 0.994 / CC1/2 anomalous: 0.026 / Rmerge(I) obs: 0.1987 / Rpim(I) all: 0.1024 / Rrim(I) all: 0.2248 / AbsDiff over sigma anomalous: 0.803 / Baniso tensor eigenvalue 1: 124.6775 Å2 / Baniso tensor eigenvalue 2: 124.6775 Å2 / Baniso tensor eigenvalue 3: 0 Å2 / Baniso tensor eigenvector 1 ortho1: 1 / Baniso tensor eigenvector 1 ortho2: 0 / Baniso tensor eigenvector 1 ortho3: 0 / Baniso tensor eigenvector 2 ortho1: 0 / Baniso tensor eigenvector 2 ortho2: 1 / Baniso tensor eigenvector 2 ortho3: 0 / Baniso tensor eigenvector 3 ortho1: 0 / Baniso tensor eigenvector 3 ortho2: 0 / Baniso tensor eigenvector 3 ortho3: 1 / Aniso diffraction limit 1: 3.96 Å / Aniso diffraction limit 2: 3.96 Å / Aniso diffraction limit 3: 2.257 Å / Aniso diffraction limit axis 1 ortho1: 1 / Aniso diffraction limit axis 1 ortho2: 0 / Aniso diffraction limit axis 1 ortho3: 0 / Aniso diffraction limit axis 2 ortho1: 0 / Aniso diffraction limit axis 2 ortho2: 1 / Aniso diffraction limit axis 2 ortho3: 0 / Aniso diffraction limit axis 3 ortho1: 0 / Aniso diffraction limit axis 3 ortho2: 0 / Aniso diffraction limit axis 3 ortho3: 1 / Net I/σ(I): 6.17 / Num. measured all: 63019 / Observed signal threshold: 1.2 / Orthogonalization convention: pdb / % possible anomalous: 88.6 / % possible ellipsoidal: 89.9 / % possible ellipsoidal anomalous: 88.6 / % possible spherical: 37.3 / % possible spherical anomalous: 33 / Redundancy anomalous: 2.61 / Signal type: local 反射 シェル |
|
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 開始モデル: Ensemble derived from PDB ID 7ZBM  7zbm 解像度: 2.7→51.147 Å / SU ML: 0.3019 / 交差検証法: FREE R-VALUE / σ(F): 1.34 / 位相誤差: 24.9194 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.1 Å / 溶媒モデル: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 25.55 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 2.7→51.147 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: X-RAY DIFFRACTION
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ |
|
