 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7z8f | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
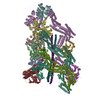
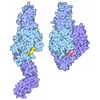
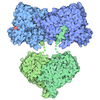
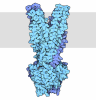
| タイトル | Composite structure of dynein-dynactin-BICDR on microtubules | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | STRUCTURAL PROTEIN / Dynein / dynactin / cargo transport / activating adaptor / cytoskeleton | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Golgi to secretory granule transport / RHOD GTPase cycle / Factors involved in megakaryocyte development and platelet production / retrograde axonal transport of mitochondrion / Regulation of actin dynamics for phagocytic cup formation / EPHB-mediated forward signaling / Adherens junctions interactions / VEGFA-VEGFR2 Pathway / Cell-extracellular matrix interactions / RHO GTPases Activate WASPs and WAVEs ...Golgi to secretory granule transport / RHOD GTPase cycle / Factors involved in megakaryocyte development and platelet production / retrograde axonal transport of mitochondrion / Regulation of actin dynamics for phagocytic cup formation / EPHB-mediated forward signaling / Adherens junctions interactions / VEGFA-VEGFR2 Pathway / Cell-extracellular matrix interactions / RHO GTPases Activate WASPs and WAVEs / MAP2K and MAPK activation / UCH proteinases / Gap junction degradation / Formation of annular gap junctions / RHOF GTPase cycle / centriolar subdistal appendage / Clathrin-mediated endocytosis / Formation of the dystrophin-glycoprotein complex (DGC) / positive regulation of neuromuscular junction development / dynactin complex / centriole-centriole cohesion / Regulation of PLK1 Activity at G2/M Transition / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Anchoring of the basal body to the plasma membrane / AURKA Activation by TPX2 / transport along microtubule / Recruitment of mitotic centrosome proteins and complexes / visual behavior / F-actin capping protein complex / WASH complex / microtubule anchoring at centrosome / dynein light chain binding / ventral spinal cord development / dynein heavy chain binding / retromer complex / cytoskeleton-dependent cytokinesis / ciliary tip / microtubule plus-end / nuclear membrane disassembly / cellular response to cytochalasin B / Intraflagellar transport / positive regulation of intracellular transport / positive regulation of microtubule nucleation / regulation of transepithelial transport / regulation of metaphase plate congression / morphogenesis of a polarized epithelium / structural constituent of postsynaptic actin cytoskeleton / positive regulation of spindle assembly / barbed-end actin filament capping / melanosome transport / protein localization to adherens junction / establishment of spindle localization / Neutrophil degranulation / dense body / postsynaptic actin cytoskeleton / Tat protein binding / coronary vasculature development / non-motile cilium assembly / regulation of cell morphogenesis / adherens junction assembly / dynein complex / retrograde transport, endosome to Golgi / COPI-independent Golgi-to-ER retrograde traffic / retrograde axonal transport / apical protein localization / RHO GTPases activate IQGAPs / RHO GTPases Activate Formins / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / MHC class II antigen presentation / Recruitment of NuMA to mitotic centrosomes / P-body assembly / microtubule motor activity / tight junction / minus-end-directed microtubule motor activity / microtubule associated complex / cytoplasmic dynein complex / dynein light intermediate chain binding / COPI-mediated anterograde transport / centrosome localization / aorta development / ventricular septum development / neuromuscular process / microtubule-based movement / nuclear migration / apical junction complex / regulation of norepinephrine uptake / transporter regulator activity / neuromuscular junction development / nitric-oxide synthase binding / cortical cytoskeleton / establishment or maintenance of cell polarity / NuA4 histone acetyltransferase complex / dynein complex binding / cell leading edge / motor behavior / dynein intermediate chain binding / cleavage furrow / brush border / establishment of mitotic spindle orientation 類似検索 - 分子機能 | ||||||||||||
| 生物種 |   Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 20 Å | ||||||||||||
 データ登録者 データ登録者 | Chaaban, S. / Carter, A.P. | ||||||||||||
| 資金援助 |  英国, European Union, 3件 英国, European Union, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2022 タイトル: Structure of dynein-dynactin on microtubules shows tandem adaptor binding. 著者: Sami Chaaban / Andrew P Carter / 要旨: Cytoplasmic dynein is a microtubule motor that is activated by its cofactor dynactin and a coiled-coil cargo adaptor. Up to two dynein dimers can be recruited per dynactin, and interactions between ...Cytoplasmic dynein is a microtubule motor that is activated by its cofactor dynactin and a coiled-coil cargo adaptor. Up to two dynein dimers can be recruited per dynactin, and interactions between them affect their combined motile behaviour. Different coiled-coil adaptors are linked to different cargos, and some share motifs known to contact sites on dynein and dynactin. There is limited structural information on how the resulting complex interacts with microtubules and how adaptors are recruited. Here we develop a cryo-electron microscopy processing pipeline to solve the high-resolution structure of dynein-dynactin and the adaptor BICDR1 bound to microtubules. This reveals the asymmetric interactions between neighbouring dynein motor domains and how they relate to motile behaviour. We found that two adaptors occupy the complex. Both adaptors make similar interactions with the dyneins but diverge in their contacts with each other and dynactin. Our structure has implications for the stability and stoichiometry of motor recruitment by cargos. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7z8f.cif.gz | 3.7 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7z8f.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 7z8f.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7z8f_validation.pdf.gz | 2.3 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7z8f_full_validation.pdf.gz | 2.3 MB | 表示 | |
| XML形式データ | 7z8f_validation.xml.gz | 485.7 KB | 表示 | |
| CIF形式データ | 7z8f_validation.cif.gz | 886.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/z8/7z8fftp://data.pdbj.org/pub/pdb/validation_reports/z8/7z8f | HTTPS FTP |
-関連構造データ















-リンク
 PDBj
PDBj



























- 集合体
集合体
| 登録構造単位 | 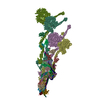
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 8種, 21分子 ABCDEFGIHJKLUWXwxklst
| #1: タンパク質 | 分子量: 42670.688 Da / 分子数: 8 / 由来タイプ: 天然 / 由来: (天然) #2: タンパク質 | | 分子量: 41782.660 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #3: タンパク質 | | 分子量: 46250.785 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #4: タンパク質 | | 分子量: 33059.848 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #5: タンパク質 | | 分子量: 30669.768 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #9: タンパク質 | | 分子量: 20703.910 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #11: タンパク質 | 分子量: 65377.035 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) 発現宿主:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: A0JNT9 #16: タンパク質 | 分子量: 10934.576 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: DYNLRB1, BITH, DNCL2A, DNLC2A, ROBLD1, HSPC162発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9NP97 |
|---|
-Dynactin subunit ... , 5種, 10分子 MNPQORSTVY
| #6: タンパク質 | 分子量: 44704.414 Da / 分子数: 4 / 由来タイプ: 天然 / 由来: (天然) #7: タンパク質 | 分子量: 21192.477 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #8: タンパク質 | 分子量: 142015.484 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #10: タンパク質 | | 分子量: 20150.533 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #12: タンパク質 | | 分子量: 52920.434 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|
-Cytoplasmic dynein 1 ... , 3種, 12分子 efmnghopijqr
| #13: タンパク質 | 分子量: 533055.125 Da / 分子数: 4 / Mutation: R1567E, K1610E / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: DYNC1H1, DHC1, DNCH1, DNCL, DNECL, DYHC, KIAA0325 発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q14204 #14: タンパク質 | 分子量: 71546.445 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: DYNC1I2, DNCI2, DNCIC2発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q13409 #15: タンパク質 | 分子量: 54173.156 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: DYNC1LI2, DNCLI2, LIC2発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: O43237 |
|---|
-非ポリマー , 5種, 43分子 



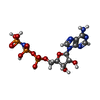
| #17: 化合物 | ChemComp-ADP /  分子量: 427.201 Da / 分子数: 13 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 13 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM#18: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 14 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 14 / 由来タイプ: 合成 / 式: Mg#19: 化合物 | ChemComp-ATP /  分子量: 507.181 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / コメント: ATP, エネルギー貯蔵分子*YM 分子量: 507.181 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / コメント: ATP, エネルギー貯蔵分子*YM#20: 化合物 |  分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn#21: 化合物 | ChemComp-ANP /  分子量: 506.196 Da / 分子数: 8 / 由来タイプ: 合成 / 式: C10H17N6O12P3 分子量: 506.196 Da / 分子数: 8 / 由来タイプ: 合成 / 式: C10H17N6O12P3コメント: AMP-PNP, エネルギー貯蔵分子類似体*YM |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 |
| |||||||||||||||||||||||||||||||||||||||||||||
| 由来(天然) |
| |||||||||||||||||||||||||||||||||||||||||||||
| 由来(組換発現) |
| |||||||||||||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.2 | |||||||||||||||||||||||||||||||||||||||||||||
| 緩衝液成分 |
| |||||||||||||||||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | |||||||||||||||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 293.15 K / 詳細: 20 second incubation |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 81000 X / 最大 デフォーカス(公称値): 4000 nm / 最小 デフォーカス(公称値): 1200 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 3 sec. / 電子線照射量: 53 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 撮影したグリッド数: 14 / 実像数: 88715 詳細: Images were collected in movie-mode and fractionated into 53 movie frames |
| 電子光学装置 | エネルギーフィルター名称: GIF Quantum LS / エネルギーフィルタースリット幅: 20 eV |
| 画像スキャン | 横: 5760 / 縦: 4092 |
- 解析
解析
| EMソフトウェア |
| |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||
| 3次元再構成 | 解像度: 20 Å / 解像度の算出法: OTHER / 粒子像の数: 628033 詳細: This is a composite of multiple maps with resolutions ranging from 3.3-12.2 A, resampled on a grid of 2.5 A/pix 対称性のタイプ: POINT | |||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: REAL |
