 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7pds | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | The structure of PriRep1 with dsDNA | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | REPLICATION / Helicase / SaPI1 | ||||||
| 機能・相同性 | Virulence-associated E / Virulence-associated protein E-like domain / P-loop containing nucleoside triphosphate hydrolase / PHOSPHOTHIOPHOSPHORIC ACID-ADENYLATE ESTER / DNA / Similar to D. nodosus vapE 機能・相同性情報 機能・相同性情報 | ||||||
| 生物種 |   Staphylococcus aureus (黄色ブドウ球菌) Staphylococcus aureus (黄色ブドウ球菌) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.14 Å | ||||||
 データ登録者 データ登録者 | Qiao, C.C. / Mir Sanchis, I. | ||||||
| 資金援助 |  スウェーデン, 1件 スウェーデン, 1件
| ||||||
 引用 引用 | ジャーナル: Nucleic Acids Res / 年: 2022 タイトル: Staphylococcal self-loading helicases couple the staircase mechanism with inter domain high flexibility. 著者: Cuncun Qiao / Gianluca Debiasi-Anders / Ignacio Mir-Sanchis / 要旨: Replication is a crucial cellular process. Replicative helicases unwind DNA providing the template strand to the polymerase and promoting replication fork progression. Helicases are multi-domain ...Replication is a crucial cellular process. Replicative helicases unwind DNA providing the template strand to the polymerase and promoting replication fork progression. Helicases are multi-domain proteins which use an ATPase domain to couple ATP hydrolysis with translocation, however the role that the other domains might have during translocation remains elusive. Here, we studied the unexplored self-loading helicases called Reps, present in Staphylococcus aureus pathogenicity islands (SaPIs). Our cryoEM structures of the PriRep5 from SaPI5 (3.3 Å), the Rep1 from SaPI1 (3.9 Å) and Rep1-DNA complex (3.1Å) showed that in both Reps, the C-terminal domain (CTD) undergoes two distinct movements respect the ATPase domain. We experimentally demonstrate both in vitro and in vivo that SaPI-encoded Reps need key amino acids involved in the staircase mechanism of translocation. Additionally, we demonstrate that the CTD's presence is necessary for the maintenance of full ATPase and helicase activities. We speculate that this high interdomain flexibility couples Rep's activities as initiators and as helicases. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7pds.cif.gz | 484.1 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7pds.ent.gz | 402.7 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7pds.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7pds_validation.pdf.gz | 1.8 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7pds_full_validation.pdf.gz | 1.8 MB | 表示 | |
| XML形式データ | 7pds_validation.xml.gz | 88.1 KB | 表示 | |
| CIF形式データ | 7pds_validation.cif.gz | 129.8 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/pd/7pdsftp://data.pdbj.org/pub/pdb/validation_reports/pd/7pds | HTTPS FTP |
-関連構造データ
| 関連構造データ |  13342MC  7olaC  7om0C M: このデータのモデリングに利用したマップデータ C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ | |
| 電子顕微鏡画像生データ | EMPIAR-10880 (タイトル: Staphylococcal self-loading helicases couple the staircase mechanism with inter domain high flexibility Data size: 395.0 / Data #1: PriRep1-DNA [micrographs - multiframe]) |




-リンク
 PDBj
PDBj
- 集合体
集合体
| 登録構造単位 | 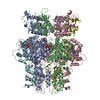
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 55423.895 Da / 分子数: 6 / 由来タイプ: 組換発現 由来: (組換発現) Staphylococcus aureus (黄色ブドウ球菌)発現宿主: #2: DNA鎖 | | 分子量: 1512.063 Da / 分子数: 1 / 由来タイプ: 合成 由来: (合成) Staphylococcus aureus (黄色ブドウ球菌)#3: 化合物 | ChemComp-AGS / 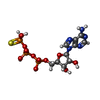  分子量: 523.247 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C10H16N5O12P3S / タイプ: SUBJECT OF INVESTIGATION 分子量: 523.247 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C10H16N5O12P3S / タイプ: SUBJECT OF INVESTIGATIONコメント: ATP-gamma-S, エネルギー貯蔵分子類似体*YM #4: 化合物 | ChemComp-MG /   分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION研究の焦点であるリガンドがあるか | Y | Has protein modification | N | |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: PriRep1-dsDNA / タイプ: COMPLEX / Entity ID: #1-#2 / 由来: RECOMBINANT |
|---|---|
| 分子量 | 値: 0.3 MDa / 実験値: YES |
| 由来(天然) | 生物種: Staphylococcus aureus (黄色ブドウ球菌) / 株: SaPI1 |
| 由来(組換発現) | 生物種: |
| 緩衝液 | pH: 8 |
| 緩衝液成分 | 濃度: 20 mM/L / 式: Tris |
| 試料 | 濃度: 0.22 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | グリッドの材料: COPPER / グリッドのタイプ: Quantifoil R2/1 |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 165000 X / 最大 デフォーカス(公称値): 3000 nm / 最小 デフォーカス(公称値): 1500 nm / Cs: 2.7 mm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 5 sec. / 電子線照射量: 1.44 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 QUANTUM (4k x 4k) 撮影したグリッド数: 16 / 実像数: 3121 |
| 画像スキャン | 動画フレーム数/画像: 40 |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: 1.18.2_3874: / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 538681 | ||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.14 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 141445 / クラス平均像の数: 2 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: AB INITIO MODEL / 空間: REAL / Target criteria: correction coefficient | ||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|
