 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7mo0 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Crystal Structure of Nucleoporin NUP50 Ran-Binding Domain in Complex with Ran-GPPNHP | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | TRANSPORT PROTEIN / nuclear pore complex component / nucleocytoplasmic transport / complex (small GTPase-nuclear protein) | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報neural tube formation / pre-miRNA export from nucleus / RNA nuclear export complex / snRNA import into nucleus / manchette / cellular response to mineralocorticoid stimulus / Nuclear Pore Complex (NPC) Disassembly / Transport of Ribonucleoproteins into the Host Nucleus / Regulation of Glucokinase by Glucokinase Regulatory Protein / Defective TPR may confer susceptibility towards thyroid papillary carcinoma (TPC) ...neural tube formation / pre-miRNA export from nucleus / RNA nuclear export complex / snRNA import into nucleus / manchette / cellular response to mineralocorticoid stimulus / Nuclear Pore Complex (NPC) Disassembly / Transport of Ribonucleoproteins into the Host Nucleus / Regulation of Glucokinase by Glucokinase Regulatory Protein / Defective TPR may confer susceptibility towards thyroid papillary carcinoma (TPC) / Regulation of cholesterol biosynthesis by SREBP (SREBF) / Transport of the SLBP independent Mature mRNA / importin-alpha family protein binding / NS1 Mediated Effects on Host Pathways / Transport of the SLBP Dependant Mature mRNA / SUMOylation of SUMOylation proteins / Transport of Mature mRNA Derived from an Intronless Transcript / protein localization to nucleolus / Rev-mediated nuclear export of HIV RNA / Nuclear import of Rev protein / SUMOylation of RNA binding proteins / NEP/NS2 Interacts with the Cellular Export Machinery / GTP metabolic process / tRNA processing in the nucleus / Transport of Mature mRNA derived from an Intron-Containing Transcript / Postmitotic nuclear pore complex (NPC) reformation / nucleocytoplasmic transport / MicroRNA (miRNA) biogenesis / Viral Messenger RNA Synthesis / DNA metabolic process / SUMOylation of ubiquitinylation proteins / Vpr-mediated nuclear import of PICs / dynein intermediate chain binding / mitotic sister chromatid segregation / SUMOylation of DNA replication proteins / ribosomal large subunit export from nucleus / spermatid development / Regulation of HSF1-mediated heat shock response / viral process / mRNA transport / positive regulation of protein binding / sperm flagellum / nuclear pore / ribosomal subunit export from nucleus / SUMOylation of DNA damage response and repair proteins / ribosomal small subunit export from nucleus / centriole / protein export from nucleus / SUMOylation of chromatin organization proteins / HCMV Late Events / mitotic spindle organization / male germ cell nucleus / hippocampus development / Transcriptional regulation by small RNAs / ISG15 antiviral mechanism / recycling endosome / positive regulation of protein import into nucleus / HCMV Early Events / protein import into nucleus / GDP binding / melanosome / nuclear envelope / mitotic cell cycle / G protein activity / actin cytoskeleton organization / snRNP Assembly / midbody / nuclear membrane / 加水分解酵素; 酸無水物に作用; GTPに作用・細胞または細胞小器官の運動に関与 / cadherin binding / protein heterodimerization activity / protein domain specific binding / cell division / GTPase activity / chromatin binding / chromatin / GTP binding / protein-containing complex binding / nucleolus / SARS-CoV-2 activates/modulates innate and adaptive immune responses / magnesium ion binding / protein-containing complex / RNA binding / extracellular exosome / nucleoplasm / nucleus / membrane / cytosol / cytoplasm 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||
| 手法 |  X線回折 / シンクロトロン / 単波長異常分散 / 解像度: 2.45 Å X線回折 / シンクロトロン / 単波長異常分散 / 解像度: 2.45 Å | ||||||||||||
 データ登録者 データ登録者 | Bley, C.J. / Nie, S. / Mobbs, G.W. / Petrovic, S. / Gres, A.T. / Liu, X. / Mukherjee, S. / Harvey, S. / Huber, F.M. / Lin, D.H. ...Bley, C.J. / Nie, S. / Mobbs, G.W. / Petrovic, S. / Gres, A.T. / Liu, X. / Mukherjee, S. / Harvey, S. / Huber, F.M. / Lin, D.H. / Brown, B. / Tang, A.W. / Rundlet, E.J. / Correia, A.R. / Chen, S. / Regmi, S.G. / Stevens, T.A. / Jette, C.A. / Dasso, M. / Patke, A. / Palazzo, A.F. / Kossiakoff, A.A. / Hoelz, A. | ||||||||||||
| 資金援助 |  米国, 3件 米国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Science / 年: 2022 タイトル: Architecture of the cytoplasmic face of the nuclear pore. 著者: Christopher J Bley / Si Nie / George W Mobbs / Stefan Petrovic / Anna T Gres / Xiaoyu Liu / Somnath Mukherjee / Sho Harvey / Ferdinand M Huber / Daniel H Lin / Bonnie Brown / Aaron W Tang / ...著者: Christopher J Bley / Si Nie / George W Mobbs / Stefan Petrovic / Anna T Gres / Xiaoyu Liu / Somnath Mukherjee / Sho Harvey / Ferdinand M Huber / Daniel H Lin / Bonnie Brown / Aaron W Tang / Emily J Rundlet / Ana R Correia / Shane Chen / Saroj G Regmi / Taylor A Stevens / Claudia A Jette / Mary Dasso / Alina Patke / Alexander F Palazzo / Anthony A Kossiakoff / André Hoelz /  要旨: INTRODUCTION The subcellular compartmentalization of eukaryotic cells requires selective transport of folded proteins and protein-nucleic acid complexes. Embedded in nuclear envelope pores, which are ...INTRODUCTION The subcellular compartmentalization of eukaryotic cells requires selective transport of folded proteins and protein-nucleic acid complexes. Embedded in nuclear envelope pores, which are generated by the circumscribed fusion of the inner and outer nuclear membranes, nuclear pore complexes (NPCs) are the sole bidirectional gateways for nucleocytoplasmic transport. The ~110-MDa human NPC is an ~1000-protein assembly that comprises multiple copies of ~34 different proteins, collectively termed nucleoporins. The symmetric core of the NPC is composed of an inner ring encircling the central transport channel and outer rings formed by Y‑shaped coat nucleoporin complexes (CNCs) anchored atop both sides of the nuclear envelope. The outer rings are decorated with compartment‑specific asymmetric nuclear basket and cytoplasmic filament nucleoporins, which establish transport directionality and provide docking sites for transport factors and the small guanosine triphosphatase Ran. The cytoplasmic filament nucleoporins also play an essential role in the irreversible remodeling of messenger ribonucleoprotein particles (mRNPs) as they exit the central transport channel. Unsurprisingly, the NPC's cytoplasmic face represents a hotspot for disease‑associated mutations and is commonly targeted by viral virulence factors. RATIONALE Previous studies established a near-atomic composite structure of the human NPC's symmetric core by combining (i) biochemical reconstitution to elucidate the interaction network between symmetric nucleoporins, (ii) crystal and single-particle cryo-electron microscopy structure determination of nucleoporins and nucleoporin complexes to reveal their three-dimensional shape and the molecular details of their interactions, (iii) quantitative docking in cryo-electron tomography (cryo-ET) maps of the intact human NPC to uncover nucleoporin stoichiometry and positioning, and (iv) cell‑based assays to validate the physiological relevance of the biochemical and structural findings. In this work, we extended our approach to the cytoplasmic filament nucleoporins to reveal the near-atomic architecture of the cytoplasmic face of the human NPC. RESULTS Using biochemical reconstitution, we elucidated the protein-protein and protein-RNA interaction networks of the human and cytoplasmic filament nucleoporins, establishing an evolutionarily conserved heterohexameric cytoplasmic filament nucleoporin complex (CFNC) held together by a central heterotrimeric coiled‑coil hub that tethers two separate mRNP‑remodeling complexes. Further biochemical analysis and determination of a series of crystal structures revealed that the metazoan‑specific cytoplasmic filament nucleoporin NUP358 is composed of 16 distinct domains, including an N‑terminal S‑shaped α‑helical solenoid followed by a coiled‑coil oligomerization element, numerous Ran‑interacting domains, an E3 ligase domain, and a C‑terminal prolyl‑isomerase domain. Physiologically validated quantitative docking into cryo-ET maps of the intact human NPC revealed that pentameric NUP358 bundles, conjoined by the oligomerization element, are anchored through their N‑terminal domains to the central stalk regions of the CNC, projecting flexibly attached domains as far as ~600 Å into the cytoplasm. Using cell‑based assays, we demonstrated that NUP358 is dispensable for the architectural integrity of the assembled interphase NPC and RNA export but is required for efficient translation. After NUP358 assignment, the remaining 4-shaped cryo‑ET density matched the dimensions of the CFNC coiled‑coil hub, in close proximity to an outer-ring NUP93. Whereas the N-terminal NUP93 assembly sensor motif anchors the properly assembled related coiled‑coil channel nucleoporin heterotrimer to the inner ring, biochemical reconstitution confirmed that the NUP93 assembly sensor is reused in anchoring the CFNC to the cytoplasmic face of the human NPC. By contrast, two CFNCs are anchored by a divergent mechanism that involves assembly sensors located in unstructured portions of two CNC nucleoporins. Whereas unassigned cryo‑ET density occupies the NUP358 and CFNC binding sites on the nuclear face, docking of the nuclear basket component ELYS established that the equivalent position on the cytoplasmic face is unoccupied, suggesting that mechanisms other than steric competition promote asymmetric distribution of nucleoporins. CONCLUSION We have substantially advanced the biochemical and structural characterization of the asymmetric nucleoporins' architecture and attachment at the cytoplasmic and nuclear faces of the NPC. Our near‑atomic composite structure of the human NPC's cytoplasmic face provides a biochemical and structural framework for elucidating the molecular basis of mRNP remodeling, viral virulence factor interference with NPC function, and the underlying mechanisms of nucleoporin diseases at the cytoplasmic face of the NPC. [Figure: see text]. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7mo0.cif.gz | 408.6 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7mo0.ent.gz | 341.3 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7mo0.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7mo0_validation.pdf.gz | 992.8 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7mo0_full_validation.pdf.gz | 1001.4 KB | 表示 | |
| XML形式データ | 7mo0_validation.xml.gz | 25.9 KB | 表示 | |
| CIF形式データ | 7mo0_validation.cif.gz | 34.8 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/mo/7mo0ftp://data.pdbj.org/pub/pdb/validation_reports/mo/7mo0 | HTTPS FTP |
-関連構造データ
| 関連構造データ |  7mniC  7mnjC  7mnkC  7mnlC  7mnmC  7mnnC  7mnoC  7mnpC  7mnqC  7mnrC  7mnsC  7mntC  7mnuC  7mnvC  7mnwC  7mnxC  7mnyC  7mnzC  7mo1C  7mo2C  7mo3C  7mo4C  7mo5C  7tblC  7tbmC C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |
-リンク
 PDBj
PDBj




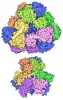







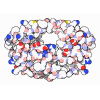







- 集合体
集合体
| 登録構造単位 | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| 単位格子 |
|
-要素
-タンパク質 , 2種, 4分子 ACBD
| #1: タンパク質 | 分子量: 24730.764 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RAN, ARA24, OK/SW-cl.81 / 発現宿主:  #2: タンパク質 | 分子量: 15221.078 Da / 分子数: 2 Fragment: RAN-binding domain of NUP50 (UNP residues 337-468) 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NUP50, NPAP60L, PRO1146 / 発現宿主: |
|---|
-非ポリマー , 4種, 68分子 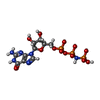


| #3: 化合物 |  分子量: 522.196 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H17N6O13P3 分子量: 522.196 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H17N6O13P3コメント: GppNHp, GMPPNP, エネルギー貯蔵分子類似体*YM #4: 化合物 |  分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg#5: 化合物 | ChemComp-SO4 / |  分子量: 96.063 Da / 分子数: 1 / 由来タイプ: 合成 / 式: SO4 分子量: 96.063 Da / 分子数: 1 / 由来タイプ: 合成 / 式: SO4#6: 水 | ChemComp-HOH / | 分子量: 18.015 Da / 分子数: 63 / 由来タイプ: 天然 / 式: H2O |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 2.35 Å3/Da / 溶媒含有率: 47.68 % |
|---|---|
| 結晶化 | 温度: 294 K / 手法: 蒸気拡散法, ハンギングドロップ法 / pH: 7.5 詳細: 25% w/v PEG3350, 0.2 M ammonium sulfate, 0.1 M HEPES |
-データ収集
| 回折 | 平均測定温度: 100 K / Serial crystal experiment: N | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: APS / ビームライン: 23-ID-D / 波長: 0.97942 Å | |||||||||||||||||||||
| 検出器 | タイプ: DECTRIS PILATUS 6M / 検出器: PIXEL / 日付: 2019年8月7日 | |||||||||||||||||||||
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray | |||||||||||||||||||||
| 放射波長 | 波長: 0.97942 Å / 相対比: 1 | |||||||||||||||||||||
| 反射 | 解像度: 2.45→29.73 Å / Num. obs: 52982 / % possible obs: 100 % / 冗長度: 13.1 % / Biso Wilson estimate: 56.22 Å2 / Rpim(I) all: 0.034 / Rrim(I) all: 0.125 / Net I/σ(I): 15.3 | |||||||||||||||||||||
| 反射 シェル | Diffraction-ID: 1 / % possible all: 100
|
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 単波長異常分散 / 解像度: 2.45→29.73 Å / SU ML: 0.33 / 交差検証法: THROUGHOUT / σ(F): 1.34 / 位相誤差: 30.01 / 立体化学のターゲット値: ML
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å / 溶媒モデル: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso max: 203.36 Å2 / Biso mean: 87.1956 Å2 / Biso min: 36.84 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: final / 解像度: 2.45→29.73 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル | Refine-ID: X-RAY DIFFRACTION / Rfactor Rfree error: 0 / Total num. of bins used: 19
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: X-RAY DIFFRACTION
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ |
|
