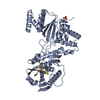
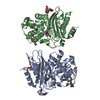
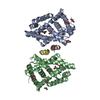
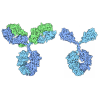
登録情報 データベース : PDB / ID : 6xyrタイトル Structure of the T4Lnano fusion protein T4Lnano,Endolysin,Calmodulin,Endolysin,Calmodulin-1 キーワード / / / / 機能・相同性 分子機能 ドメイン・相同性 構成要素
/ / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / 生物種 Homo sapiens (ヒト)手法 / / / / 解像度 : 2.079 Å データ登録者 Benoit, R.M. / Bierig, T. / Collu, C. / Engilberge, S. / Olieric, V. 資金援助 組織 認可番号 国 Novartis FreeNovation Promedica Siftung
ジャーナル : Structure / 年 : 2022タイトル : Chimeric single α-helical domains as rigid fusion protein connections for protein nanotechnology and structural biology.
著者 :
Gabriella Collu / Tobias Bierig / Anna-Sophia Krebs / Sylvain Engilberge / Niveditha Varma / Ramon Guixà-González / Timothy Sharpe / Xavier Deupi / Vincent Olieric / Emiliya Poghosyan / Roger M Benoit / 要旨 :
Chimeric fusion proteins are essential tools for protein nanotechnology. Non-optimized protein-protein connections are usually flexible and therefore unsuitable as structural building blocks. Here we ... Chimeric fusion proteins are essential tools for protein nanotechnology. Non-optimized protein-protein connections are usually flexible and therefore unsuitable as structural building blocks. Here we show that the ER/K motif, a single α-helical domain (SAH), can be seamlessly fused to terminal helices of proteins, forming an extended, partially free-standing rigid helix. This enables the connection of two domains at a defined distance and orientation. We designed three constructs termed YFPnano, T4Lnano, and MoStoNano. Analysis of experimentally determined structures and molecular dynamics simulations reveals a certain degree of plasticity in the connections that allows the adaptation to crystal contact opportunities. Our data show that SAHs can be stably integrated into designed structural elements, enabling new possibilities for protein nanotechnology, for example, to improve the exposure of epitopes on nanoparticles (structural vaccinology), to engineer crystal contacts with minimal impact on construct flexibility (for the study of protein dynamics), and to design novel biomaterials. #1: ジャーナル : Biorxiv / 年 : 2020タイトル : Chimeric single alpha-helical domains as rigid fusion protein connections for protein nanotechnology and structural biology
著者 :
Collu, G. / Bierig, T. / Krebs, A.-S. / Engilberge, S. / Varma, N. / Guixa-Gonzalez, R. / Deupi, X. / Olieric, V. / Poghosyan, E. / Benoit, R.M. 履歴 登録 2020年1月31日 登録サイト / 処理サイト 改定 1.0 2020年12月9日 Provider / タイプ 改定 1.1 2021年10月13日 Group / Data collection / Database referencesカテゴリ citation / citation_author ... citation / citation_author / database_2 / pdbx_database_proc / pdbx_unobs_or_zero_occ_atoms Item / _database_2.pdbx_database_accession改定 1.2 2022年1月19日 Group / カテゴリ Item / _citation.page_first / _citation.year改定 1.3 2024年6月19日 Group / Database references / カテゴリ / chem_comp_bond / citation / Item
すべて表示 表示を減らす
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) Enterobacteria phage T4 (ファージ)
Enterobacteria phage T4 (ファージ) X線回折 /
X線回折 /  データ登録者
データ登録者 スイス, 2件
スイス, 2件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード






 PDBj
PDBj




















 集合体
集合体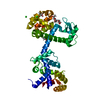


 分子量: 40.078 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Ca
分子量: 40.078 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Ca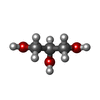
 分子量: 92.094 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C3H8O3
分子量: 92.094 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C3H8O3
 分子量: 35.453 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Cl
分子量: 35.453 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Cl 分子量: 18.015 Da / 分子数: 135 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 135 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 解析
解析