 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6u59 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | HIV-1 B41 SOSIP.664 in complex with rabbit antibody 13B | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | VIRAL PROTEIN/IMMUNE SYSTEM / HIV-1 / rabbit antibody / SOSIP / VIRAL PROTEIN-IMMUNE SYSTEM complex | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報positive regulation of plasma membrane raft polarization / positive regulation of receptor clustering / host cell endosome membrane / clathrin-dependent endocytosis of virus by host cell / viral protein processing / fusion of virus membrane with host plasma membrane / fusion of virus membrane with host endosome membrane / viral envelope / virion attachment to host cell / host cell plasma membrane ...positive regulation of plasma membrane raft polarization / positive regulation of receptor clustering / host cell endosome membrane / clathrin-dependent endocytosis of virus by host cell / viral protein processing / fusion of virus membrane with host plasma membrane / fusion of virus membrane with host endosome membrane / viral envelope / virion attachment to host cell / host cell plasma membrane / virion membrane / structural molecule activity / membrane 類似検索 - 分子機能 | ||||||||||||
| 生物種 |   Human immunodeficiency virus 1 (ヒト免疫不全ウイルス) Human immunodeficiency virus 1 (ヒト免疫不全ウイルス) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.86 Å | ||||||||||||
 データ登録者 データ登録者 | Yang, Y.R. / Ward, A.B. | ||||||||||||
| 資金援助 |  米国, 3件 米国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: J Virol / 年: 2020 タイトル: Autologous Antibody Responses to an HIV Envelope Glycan Hole Are Not Easily Broadened in Rabbits. 著者: Yuhe R Yang / Laura E McCoy / Marit J van Gils / Raiees Andrabi / Hannah L Turner / Meng Yuan / Christopher A Cottrell / Gabriel Ozorowski / James Voss / Matthias Pauthner / Thomas M ...著者: Yuhe R Yang / Laura E McCoy / Marit J van Gils / Raiees Andrabi / Hannah L Turner / Meng Yuan / Christopher A Cottrell / Gabriel Ozorowski / James Voss / Matthias Pauthner / Thomas M Polveroni / Terrence Messmer / Ian A Wilson / Rogier W Sanders / Dennis R Burton / Andrew B Ward /   要旨: Extensive studies with subtype A BG505-derived HIV envelope glycoprotein (Env) immunogens have revealed that the dominant autologous neutralizing epitope in rabbits is located in an exposed region of ...Extensive studies with subtype A BG505-derived HIV envelope glycoprotein (Env) immunogens have revealed that the dominant autologous neutralizing epitope in rabbits is located in an exposed region of the heavily glycosylated trimer that lacks potential N-linked glycosylation sites at positions 230, 241, and 289. The Env derived from B41, a subtype B virus, shares a glycan hole centered on positions 230 and 289. To test whether broader neutralization to the common glycan hole can be achieved, we immunized rabbits with B41 SOSIP (gp120-gp41 disulfide [SOS] with an isoleucine-to-proline mutation [IP] in gp41) alone, as well as B41 and BG505 coimmunization. We isolated autologous neutralizing antibodies (nAbs) and described their structure in complex with the B41 Env. Our data suggest that distinct autologous nAb lineages are induced by BG505 and B41 immunogens, even when both were administered together. In contrast to previously described BG505 glycan hole antibodies, the B41-specific nAbs accommodate the >97% conserved N241 glycan, which is present in B41. Single-particle cryo-electron microscopy studies confirmed that B41- and BG505-specific nAbs bind to overlapping glycan hole epitopes. We then used our high-resolution data to guide mutations in the BG505 glycan hole epitope in an attempt to broaden the reactivity of a B41-specific nAb, but we recovered only partial binding. Our data demonstrate that the lack of cross-reactivity in glycan hole antibodies is due to amino acid differences within the epitope, and our attempts to rationally design cross-reactive trimers resulted in only limited success. Thus, even for the immunodominant glycan hole shared between BG505 and B41, the prospect of designing prime-boost immunogens remains difficult. A glycan hole is one of the most dominant autologous neutralizing epitopes targeted on BG505 and B41 SOSIP trimer-immunized rabbits. Our high-resolution cryo-electron microscopy (cryoEM) studies of B41 in complex with a B41-specific antibody complex elucidate the molecular basis of this strain-specific glycan hole response. We conclude that even for the immunodominant glycan hole shared between BG505 and B41, the prospect of designing prime-boost immunogens remains difficult. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6u59.cif.gz | 518.7 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6u59.ent.gz | 422.2 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6u59.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 6u59_validation.pdf.gz | 3.6 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 6u59_full_validation.pdf.gz | 3.7 MB | 表示 | |
| XML形式データ | 6u59_validation.xml.gz | 78.6 KB | 表示 | |
| CIF形式データ | 6u59_validation.cif.gz | 119.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/u5/6u59ftp://data.pdbj.org/pub/pdb/validation_reports/u5/6u59 | HTTPS FTP |
-関連構造データ




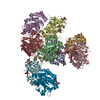
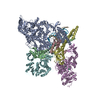
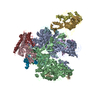
-リンク
 PDBj
PDBj

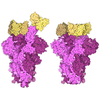
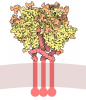
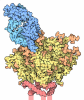

- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 2種, 6分子 ACGBDI
| #1: タンパク質 | 分子量: 58872.902 Da / 分子数: 3 / 由来タイプ: 組換発現 由来: (組換発現) Human immunodeficiency virus 1 (ヒト免疫不全ウイルス)株: B41 / 細胞株 (発現宿主): HEK 293F / 発現宿主:  Homo sapiens (ヒト) / 参照: UniProt: B3UES2 Homo sapiens (ヒト) / 参照: UniProt: B3UES2#2: タンパク質 | 分子量: 17357.824 Da / 分子数: 3 / 由来タイプ: 組換発現 由来: (組換発現) Human immunodeficiency virus 1 (ヒト免疫不全ウイルス)株: B41 / 細胞株 (発現宿主): HEK 293F / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: B3UEZ6 |
|---|
-抗体 , 2種, 6分子 LEJHFK
| #3: 抗体 | 分子量: 11977.202 Da / 分子数: 3 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)#4: 抗体 | 分子量: 12790.192 Da / 分子数: 3 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) |
|---|
-糖 , 4種, 69分子 
| #5: 多糖 | 2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose #6: 多糖 | #7: 多糖 | #8: 糖 | ChemComp-NAG /  タイプ: D-saccharide, beta linking / 分子量: 221.208 Da / 分子数: 30 / 由来タイプ: 組換発現 / 式: C8H15NO6 タイプ: D-saccharide, beta linking / 分子量: 221.208 Da / 分子数: 30 / 由来タイプ: 組換発現 / 式: C8H15NO6 |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.57 MDa / 実験値: NO | ||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||
| 緩衝液 | pH: 7.4 詳細: Detergent was added to sample shortly prior to freezing. | ||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||
| 試料 | 濃度: 5 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 400 divisions/in. / グリッドのタイプ: C-flat-2/2 | ||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Talos Arctica / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TALOS ARCTICA |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 36000 X / 最大 デフォーカス(公称値): 2000 nm / 最小 デフォーカス(公称値): 500 nm / Cs: 2.7 mm / C2レンズ絞り径: 70 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 14 sec. / 電子線照射量: 60 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 1 / 実像数: 1721 |
| 画像スキャン | サンプリングサイズ: 5 µm / 動画フレーム数/画像: 56 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C3 (3回回転対称) | ||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.86 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 147520 / クラス平均像の数: 1 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | 空間: REAL |
