ジャーナル: Structure / 年: 2022 タイトル: Chimeric single α-helical domains as rigid fusion protein connections for protein nanotechnology and structural biology. 著者: Gabriella Collu / Tobias Bierig / Anna-Sophia Krebs / Sylvain Engilberge / Niveditha Varma / Ramon Guixà-González / Timothy Sharpe / Xavier Deupi / Vincent Olieric / Emiliya Poghosyan / Roger M Benoit / 要旨: Chimeric fusion proteins are essential tools for protein nanotechnology. Non-optimized protein-protein connections are usually flexible and therefore unsuitable as structural building blocks. Here we ...Chimeric fusion proteins are essential tools for protein nanotechnology. Non-optimized protein-protein connections are usually flexible and therefore unsuitable as structural building blocks. Here we show that the ER/K motif, a single α-helical domain (SAH), can be seamlessly fused to terminal helices of proteins, forming an extended, partially free-standing rigid helix. This enables the connection of two domains at a defined distance and orientation. We designed three constructs termed YFPnano, T4Lnano, and MoStoNano. Analysis of experimentally determined structures and molecular dynamics simulations reveals a certain degree of plasticity in the connections that allows the adaptation to crystal contact opportunities. Our data show that SAHs can be stably integrated into designed structural elements, enabling new possibilities for protein nanotechnology, for example, to improve the exposure of epitopes on nanoparticles (structural vaccinology), to engineer crystal contacts with minimal impact on construct flexibility (for the study of protein dynamics), and to design novel biomaterials.
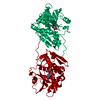
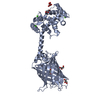
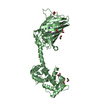
根拠: The fusion protein is a monomer. There is only an intramolecular interaction between the C-terminal Calmodulin and the N-terminal Calmodulin-binding peptide in the construct.
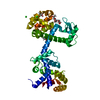
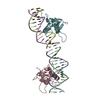
分子量: 49199.820 Da / 分子数: 2 / 由来タイプ: 組換発現 詳細: residues -35 to -32 = leftovers from cleaved-off tag res -31 t -11 = Calmodulin-binding peptide (very similar to the peptide in PDB entry 2BBM) res -10 to 0 = ER/K helix (small fragment from ...詳細: residues -35 to -32 = leftovers from cleaved-off tag res -31 t -11 = Calmodulin-binding peptide (very similar to the peptide in PDB entry 2BBM) res -10 to 0 = ER/K helix (small fragment from the protein in PDB entry 5HMO, res 818-828) res 1 to 238 = Yellow Fluorescent Protein (nearly identical to pdb entry 3V3D) res 239 - 252 = flexible linker res 253 - 400 = Calmodulin (as in PDB entry 2BBM),residues -35 to -32 = leftovers from cleaved-off tag 由来: (組換発現) Oryctolagus cuniculus (ウサギ), (組換発現) Bos taurus (ウシ), (組換発現) Aequorea victoria (オワンクラゲ), (組換発現) Homo sapiens (ヒト) 遺伝子: MYLK2, MYO10, GFP, CALM1, CALM, CAM, CAM1 / プラスミド: pET28a / 発現宿主: Escherichia coli BL21(DE3) (大腸菌) / Variant (発現宿主): Rosetta 2 参照: UniProt: P07313, UniProt: P79114, UniProt: P42212, UniProt: P0DP23, myosin-light-chain kinase
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報

 Aequorea victoria (オワンクラゲ)
Aequorea victoria (オワンクラゲ) Homo sapiens (ヒト)
Homo sapiens (ヒト) X線回折 /
X線回折 /  データ登録者
データ登録者 スイス, 2件
スイス, 2件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード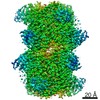








 PDBj
PDBj























 集合体
集合体

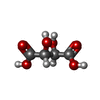

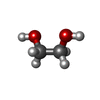
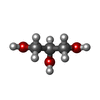


 分子量: 150.087 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C4H6O6
分子量: 150.087 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C4H6O6 分子量: 40.078 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Ca
分子量: 40.078 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Ca 分子量: 62.068 Da / 分子数: 9 / 由来タイプ: 合成 / 式: C2H6O2
分子量: 62.068 Da / 分子数: 9 / 由来タイプ: 合成 / 式: C2H6O2 分子量: 92.094 Da / 分子数: 5 / 由来タイプ: 天然 / 式: C3H8O3
分子量: 92.094 Da / 分子数: 5 / 由来タイプ: 天然 / 式: C3H8O3 分子量: 22.990 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Na
分子量: 22.990 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Na 試料調製
試料調製 解析
解析