National Institutes of Health/Office of the Director
AG-04812
United States
National Science Foundation (NSF, United States)
MCB-0958111
United States
Citation
Journal: Science / Year: 2018 Title: Atomic structures of low-complexity protein segments reveal kinked β sheets that assemble networks. Authors: Michael P Hughes / Michael R Sawaya / David R Boyer / Lukasz Goldschmidt / Jose A Rodriguez / Duilio Cascio / Lisa Chong / Tamir Gonen / David S Eisenberg / Abstract: Subcellular membraneless assemblies are a reinvigorated area of study in biology, with spirited scientific discussions on the forces between the low-complexity protein domains within these assemblies. ...Subcellular membraneless assemblies are a reinvigorated area of study in biology, with spirited scientific discussions on the forces between the low-complexity protein domains within these assemblies. To illuminate these forces, we determined the atomic structures of five segments from protein low-complexity domains associated with membraneless assemblies. Their common structural feature is the stacking of segments into kinked β sheets that pair into protofilaments. Unlike steric zippers of amyloid fibrils, the kinked sheets interact weakly through polar atoms and aromatic side chains. By computationally threading the human proteome on our kinked structures, we identified hundreds of low-complexity segments potentially capable of forming such interactions. These segments are found in proteins as diverse as RNA binders, nuclear pore proteins, and keratins, which are known to form networks and localize to membraneless assemblies.
History
Deposition
Dec 24, 2017
Deposition site: RCSB / Processing site: RCSB
Revision 1.0
Apr 4, 2018
Provider: repository / Type: Initial release
Revision 1.1
Apr 25, 2018
Group: Data collection / Category: diffrn_source / Item: _diffrn_source.source
Mass: 18.015 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Formula: H2O
-
Experimental details
-
Experiment
Experiment
Method: ELECTRON CRYSTALLOGRAPHY
EM experiment
Aggregation state: 3D ARRAY / 3D reconstruction method: electron crystallography
-
Sample preparation
Component
Name: A crystal containing protofilaments of an 8-residue segment of Nup98 Type: COMPLEX / Entity ID: #1 / Source: NATURAL
Molecular weight
Experimental value: NO
Source (natural)
Organism: Homo sapiens (human)
EM crystal formation
Instrument: hanging drop vapor diffusion / Atmosphere: air Details: The peptide was solubilized by adding nano-pure H2O with 20% DMSO to achieve a concentration of 12 mg/mL. The peptide solution was immediately used for crystallization. Crystals grew at room ...Details: The peptide was solubilized by adding nano-pure H2O with 20% DMSO to achieve a concentration of 12 mg/mL. The peptide solution was immediately used for crystallization. Crystals grew at room temperature by hanging drop vapor diffusion. Lipid mixture: none / Temperature: 298 K
Buffer solution
pH: 9.5
Buffer component
ID
Conc.
Name
Formula
Buffer-ID
1
0.1M
CHES
C8H17NO3S
1
2
10percent (v/v)
ethanol
C2H6O
1
Specimen
Conc.: 12 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES / Details: crystal
Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE
Crystal
Density Matthews: 1.45 Å3/Da / Density % sol: 15.41 %
Crystal grow
Temperature: 298 K / Method: vapor diffusion, hanging drop / pH: 9.5 / Details: 0.1 M CHES, pH 9.5, 10% ethanol
-
Data collection
Microscopy
Model: FEI TECNAI 20
Electron gun
Electron source: FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM
Electron lens
Mode: DIFFRACTION / Alignment procedure: BASIC
Specimen holder
Cryogen: NITROGEN Specimen holder model: GATAN 626 SINGLE TILT LIQUID NITROGEN CRYO TRANSFER HOLDER Temperature (max): 100 K / Temperature (min): 100 K
Image recording
Average exposure time: 2 sec. / Electron dose: 0.01 e/Å2 / Film or detector model: TVIPS TEMCAM-F415 (4k x 4k) / Num. of diffraction images: 398 / Num. of grids imaged: 2 Details: The detector was operated in rolling shutter mode with 2x2 pixel binning.
Details: Phase statistics are not applicable. No imaging was used. The phases wwere obtained by a crystallographic direct methods program, SHELXD. Fourier space coverage: 88 % / High resolution: 0.9 Å / Num. of intensities measured: 31674 / Num. of structure factors: 5800 / Phase error: 42.6 ° / Phase residual: 42.6 ° / Phase error rejection criteria: 0 / Rmerge: 0.289 / Rsym: 0.289
Diffraction
Mean temperature: 100 K
Diffraction source
Source: ELECTRON MICROSCOPE / Type: TECNAI F20 TEM / Wavelength: 0.0251 Å
Detector
Type: TVIPS F416 CMOS CAMERA / Detector: CMOS / Date: Mar 23, 2017
∠α: 93.14 ° / ∠β: 92.73 ° / ∠γ: 97.15 ° / A: 4.79 Å / B: 18.24 Å / C: 26.44 Å / Space group name: P1 / Space group num: 1
CTF correction
Type: NONE
3D reconstruction
Resolution: 0.9 Å / Resolution method: DIFFRACTION PATTERN/LAYERLINES Details: Density map was obtained using measured diffraction intensities and the phases acquired from a crystallographic direct methods program, SHELXD. Symmetry type: 3D CRYSTAL
Atomic model building
B value: 8 / Protocol: OTHER / Space: RECIPROCAL / Target criteria: maximum likelihood
Refinement
Method to determine structure: AB INITIO PHASING / Resolution: 0.9→26.36 Å / Cor.coef. Fo:Fc: 0.911 / Cor.coef. Fo:Fc free: 0.895 / SU R Cruickshank DPI: 0.032 / Cross valid method: THROUGHOUT / σ(F): 0 / SU R Blow DPI: 0.03 / SU Rfree Blow DPI: 0.033 / SU Rfree Cruickshank DPI: 0.035
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller

 Yorodumi
Yorodumi Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information Homo sapiens (human)
Homo sapiens (human) Authors
Authors United States, 3items
United States, 3items  Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads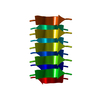










 PDBj
PDBj


















 Assembly
Assembly

 Mass: 18.015 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Formula: H2O
Mass: 18.015 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Formula: H2O Sample preparation
Sample preparation FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM
FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM Processing
Processing