 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6bj3 | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | TCR55 in complex with HIV(Pol448-456)/HLA-B35 | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | IMMUNE SYSTEM / non-agonist / complex | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報regulation of interleukin-12 production / regulation of dendritic cell differentiation / integrase activity / regulation of T cell anergy / regulation of interleukin-6 production / Integration of viral DNA into host genomic DNA / Autointegration results in viral DNA circles / T cell receptor complex / Minus-strand DNA synthesis / Plus-strand DNA synthesis ...regulation of interleukin-12 production / regulation of dendritic cell differentiation / integrase activity / regulation of T cell anergy / regulation of interleukin-6 production / Integration of viral DNA into host genomic DNA / Autointegration results in viral DNA circles / T cell receptor complex / Minus-strand DNA synthesis / Plus-strand DNA synthesis / Uncoating of the HIV Virion / 2-LTR circle formation / Vpr-mediated nuclear import of PICs / Early Phase of HIV Life Cycle / Integration of provirus / APOBEC3G mediated resistance to HIV-1 infection / protection from natural killer cell mediated cytotoxicity / TAP binding / Binding and entry of HIV virion / immune system process / detection of bacterium / antigen processing and presentation of endogenous peptide antigen via MHC class Ib / antigen processing and presentation of endogenous peptide antigen via MHC class I via ER pathway, TAP-independent / viral life cycle / secretory granule membrane / early endosome lumen / Nef mediated downregulation of MHC class I complex cell surface expression / DAP12 interactions / Endosomal/Vacuolar pathway / T cell mediated cytotoxicity / HIV-1 retropepsin / symbiont-mediated activation of host apoptosis / Antigen Presentation: Folding, assembly and peptide loading of class I MHC / retroviral ribonuclease H / exoribonuclease H / lumenal side of endoplasmic reticulum membrane / regulation of iron ion transport / cellular response to iron(III) ion / negative regulation of iron ion transport / negative regulation of forebrain neuron differentiation / antigen processing and presentation of exogenous protein antigen via MHC class Ib, TAP-dependent / exoribonuclease H activity / ER to Golgi transport vesicle membrane / peptide antigen assembly with MHC class I protein complex / regulation of erythrocyte differentiation / response to molecule of bacterial origin / Assembly Of The HIV Virion / HFE-transferrin receptor complex / defense response / transferrin transport / MHC class I peptide loading complex / cellular response to iron ion / negative regulation of receptor-mediated endocytosis / positive regulation of T cell cytokine production / antigen processing and presentation of endogenous peptide antigen via MHC class I / Budding and maturation of HIV virion / protein processing / MHC class I protein complex / peptide antigen assembly with MHC class II protein complex / viral genome integration into host DNA / negative regulation of neurogenesis / cellular response to nicotine / MHC class II protein complex / positive regulation of receptor-mediated endocytosis / establishment of integrated proviral latency / RNA-directed DNA polymerase / multicellular organismal-level iron ion homeostasis / positive regulation of T cell mediated cytotoxicity / RNA stem-loop binding / viral penetration into host nucleus / specific granule lumen / positive regulation of immune response / antigen processing and presentation of exogenous peptide antigen via MHC class II / host multivesicular body / peptide antigen binding / phagocytic vesicle membrane / RNA-directed DNA polymerase activity / recycling endosome membrane / RNA-DNA hybrid ribonuclease activity / positive regulation of T cell activation / negative regulation of epithelial cell proliferation / Interferon gamma signaling / Immunoregulatory interactions between a Lymphoid and a non-Lymphoid cell / 転移酵素; リンを含む基を移すもの; 核酸を移すもの / Interferon alpha/beta signaling / Modulation by Mtb of host immune system / positive regulation of cellular senescence / sensory perception of smell / tertiary granule lumen / MHC class II protein complex binding / DAP12 signaling / T cell differentiation in thymus / peptidase activity / late endosome membrane / host cell / negative regulation of neuron projection development / ER-Phagosome pathway / protein refolding / protein-folding chaperone binding / viral nucleocapsid 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト)  Human immunodeficiency virus 1 (ヒト免疫不全ウイルス) Human immunodeficiency virus 1 (ヒト免疫不全ウイルス) | ||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 1.898 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 1.898 Å | ||||||
 データ登録者 データ登録者 | Jude, K.M. / Sibener, L.V. / Garcia, K.C. | ||||||
| 資金援助 |  米国, 1件 米国, 1件
| ||||||
 引用 引用 | ジャーナル: Cell / 年: 2018 タイトル: Isolation of a Structural Mechanism for Uncoupling T Cell Receptor Signaling from Peptide-MHC Binding. 著者: Sibener, L.V. / Fernandes, R.A. / Kolawole, E.M. / Carbone, C.B. / Liu, F. / McAffee, D. / Birnbaum, M.E. / Yang, X. / Su, L.F. / Yu, W. / Dong, S. / Gee, M.H. / Jude, K.M. / Davis, M.M. / ...著者: Sibener, L.V. / Fernandes, R.A. / Kolawole, E.M. / Carbone, C.B. / Liu, F. / McAffee, D. / Birnbaum, M.E. / Yang, X. / Su, L.F. / Yu, W. / Dong, S. / Gee, M.H. / Jude, K.M. / Davis, M.M. / Groves, J.T. / Goddard III, W.A. / Heath, J.R. / Evavold, B.D. / Vale, R.D. / Garcia, K.C. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6bj3.cif.gz | 355.5 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6bj3.ent.gz | 289 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6bj3.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/bj/6bj3ftp://data.pdbj.org/pub/pdb/validation_reports/bj/6bj3 | HTTPS FTP |
|---|
-関連構造データ
| 関連構造データ |  6bj2C  6bj8C  1a1nS  4lfhS  4pj5S C: 同じ文献を引用 ( S: 精密化の開始モデル |
|---|---|
| 類似構造データ | |
| 実験データセット #1 | データ参照: 10.15785/SBGRID/519 / データの種類: diffraction image data / 詳細: SBGrid Data Bank |















-リンク
 PDBj
PDBj




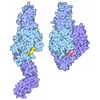


- 集合体
集合体
| 登録構造単位 | 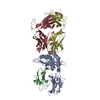
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| 単位格子 |
|
-要素
-タンパク質 , 4種, 4分子 ABDH
| #1: タンパク質 | 分子量: 31940.246 Da / 分子数: 1 / 断片: UNP residues 25-300 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: HLA-B, HLAB / 発現宿主:  Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: P30685, UniProt: P01889*PLUS Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: P30685, UniProt: P01889*PLUS |
|---|---|
| #2: タンパク質 | 分子量: 11748.160 Da / 分子数: 1 / 断片: UNP residues 21-119 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: B2M, CDABP0092, HDCMA22P / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: P61769 |
| #3: タンパク質 | 分子量: 22890.752 Da / 分子数: 1 / Mutation: T161C / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 発現宿主:  |
| #4: タンパク質 | 分子量: 27390.535 Da / 分子数: 1 / Mutation: A187C / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 発現宿主: |
-タンパク質・ペプチド , 1種, 1分子 C
| #5: タンパク質・ペプチド | 分子量: 1014.126 Da / 分子数: 1 / 由来タイプ: 合成 由来: (合成) Human immunodeficiency virus 1 (ヒト免疫不全ウイルス)参照: UniProt: P04585*PLUS |
|---|
-非ポリマー , 3種, 372分子 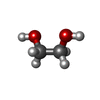

| #6: 化合物 |  分子量: 62.068 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C2H6O2 分子量: 62.068 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C2H6O2#7: 化合物 | ChemComp-SO4 /  分子量: 96.063 Da / 分子数: 5 / 由来タイプ: 合成 / 式: SO4 分子量: 96.063 Da / 分子数: 5 / 由来タイプ: 合成 / 式: SO4#8: 水 | ChemComp-HOH / | 分子量: 18.015 Da / 分子数: 364 / 由来タイプ: 天然 / 式: H2O |
|---|
-詳細
| Has protein modification | Y |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 2.87 Å3/Da / 溶媒含有率: 57.09 % |
|---|---|
| 結晶化 | 温度: 295 K / 手法: 蒸気拡散法 詳細: PEG4000, 0.1 M sodium cacodylate, pH 6.0, 0.2 M ammonium sulfate |
-データ収集
| 回折 | 平均測定温度: 100 K |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: ALS / ビームライン: 8.2.2 / 波長: 0.99995 Å |
| 検出器 | タイプ: ADSC QUANTUM 315r / 検出器: CCD / 日付: 2016年4月29日 |
| 放射 | モノクロメーター: double crystal Si(111) / プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 0.99995 Å / 相対比: 1 |
| 反射 | 解像度: 1.898→50 Å / Num. obs: 83485 / % possible obs: 97.79 % / 冗長度: 3.4 % / Biso Wilson estimate: 34.9 Å2 / CC1/2: 0.985 / Rmerge(I) obs: 0.1382 / Rpim(I) all: 0.0853 / Rrim(I) all: 0.1628 / Net I/σ(I): 4.67 |
| 反射 シェル | 解像度: 1.898→1.966 Å / 冗長度: 3.3 % / Rmerge(I) obs: 1.313 / Mean I/σ(I) obs: 0.7 / Num. unique obs: 7910 / CC1/2: 0.406 / Rpim(I) all: 0.8382 / Rrim(I) all: 1.563 / % possible all: 92.69 |
- 解析
解析
| ソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 開始モデル: PDB entries 1A1N, 4LFH, 4PJ5 解像度: 1.898→48.094 Å / SU ML: 0.29 / 交差検証法: FREE R-VALUE / σ(F): 1.33 / 位相誤差: 24.16
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 1.898→48.094 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ |
|