 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-5v7x: Crystal Structure of Myosin 1b residues 1-728 with bound sulfate ... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5v7x | ||||||
|---|---|---|---|---|---|---|---|
| Title | Crystal Structure of Myosin 1b residues 1-728 with bound sulfate and Calmodulin | ||||||
 Components Components |
| ||||||
 Keywords Keywords | MOTOR PROTEIN / myosin / motor / calmodulin / unconventional / actin / force | ||||||
| Function / homology |  Function and homology information Function and homology informationpost-Golgi vesicle-mediated transport / actin filament-based movement / myosin complex / CaM pathway / Cam-PDE 1 activation / Sodium/Calcium exchangers / Calmodulin induced events / Reduction of cytosolic Ca++ levels / Activation of Ca-permeable Kainate Receptor / CREB1 phosphorylation through the activation of CaMKII/CaMKK/CaMKIV cascasde ...post-Golgi vesicle-mediated transport / actin filament-based movement / myosin complex / CaM pathway / Cam-PDE 1 activation / Sodium/Calcium exchangers / Calmodulin induced events / Reduction of cytosolic Ca++ levels / Activation of Ca-permeable Kainate Receptor / CREB1 phosphorylation through the activation of CaMKII/CaMKK/CaMKIV cascasde / Loss of phosphorylation of MECP2 at T308 / CREB1 phosphorylation through the activation of Adenylate Cyclase / PKA activation / CaMK IV-mediated phosphorylation of CREB / Glycogen breakdown (glycogenolysis) / negative regulation of ryanodine-sensitive calcium-release channel activity / Activation of RAC1 downstream of NMDARs / microfilament motor activity / organelle localization by membrane tethering / CLEC7A (Dectin-1) induces NFAT activation / : / autophagosome membrane docking / negative regulation of high voltage-gated calcium channel activity / negative regulation of calcium ion export across plasma membrane / regulation of cardiac muscle cell action potential / presynaptic endocytosis / Synthesis of IP3 and IP4 in the cytosol / microvillus / Phase 0 - rapid depolarisation / Negative regulation of NMDA receptor-mediated neuronal transmission / phosphatidylinositol-3,4,5-trisphosphate binding / Unblocking of NMDA receptors, glutamate binding and activation / calcineurin-mediated signaling / cytoskeletal motor activity / RHO GTPases activate PAKs / regulation of cell communication by electrical coupling involved in cardiac conduction / Ion transport by P-type ATPases / brush border / Uptake and function of anthrax toxins / protein phosphatase activator activity / actin filament bundle assembly / regulation of ryanodine-sensitive calcium-release channel activity / Long-term potentiation / Calcineurin activates NFAT / Regulation of MECP2 expression and activity / DARPP-32 events / Smooth Muscle Contraction / catalytic complex / detection of calcium ion / regulation of cardiac muscle contraction / cellular response to interferon-beta / RHO GTPases activate IQGAPs / calcium channel inhibitor activity / presynaptic cytosol / Activation of AMPK downstream of NMDARs / regulation of release of sequestered calcium ion into cytosol by sarcoplasmic reticulum / eNOS activation / Ion homeostasis / regulation of calcium-mediated signaling / Tetrahydrobiopterin (BH4) synthesis, recycling, salvage and regulation / Protein methylation / titin binding / regulation of cardiac muscle contraction by regulation of the release of sequestered calcium ion / phosphatidylinositol-4,5-bisphosphate binding / voltage-gated potassium channel complex / FCERI mediated Ca+2 mobilization / calcium channel complex / substantia nigra development / regulation of heart rate / FCGR3A-mediated IL10 synthesis / calyx of Held / Antigen activates B Cell Receptor (BCR) leading to generation of second messengers / Ras activation upon Ca2+ influx through NMDA receptor / adenylate cyclase activator activity / actin filament organization / trans-Golgi network membrane / protein serine/threonine kinase activator activity / VEGFR2 mediated vascular permeability / regulation of cytokinesis / VEGFR2 mediated cell proliferation / cell periphery / positive regulation of receptor signaling pathway via JAK-STAT / spindle microtubule / sarcomere / Translocation of SLC2A4 (GLUT4) to the plasma membrane / actin filament / filopodium / calcium channel regulator activity / Transcriptional activation of mitochondrial biogenesis / transferrin transport / cellular response to type II interferon / response to calcium ion / RAF activation / G2/M transition of mitotic cell cycle / long-term synaptic potentiation / Stimuli-sensing channels / endocytosis / apical part of cell / spindle pole / calcium-dependent protein binding Similarity search - Function | ||||||
| Biological species |   Homo sapiens (human) Homo sapiens (human) | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / Resolution: 3.141 Å X-RAY DIFFRACTION / SYNCHROTRON / Resolution: 3.141 Å | ||||||
 Authors Authors | Zwolak, A. / Shuman, H. / Dominguez, R. / Ostap, E.M. | ||||||
 Citation Citation | Journal: Proc Natl Acad Sci U S A / Year: 2018 Title: High-resolution cryo-EM structures of actin-bound myosin states reveal the mechanism of myosin force sensing. Authors: Ahmet Mentes / Andrew Huehn / Xueqi Liu / Adam Zwolak / Roberto Dominguez / Henry Shuman / E Michael Ostap / Charles V Sindelar /  Abstract: Myosins adjust their power outputs in response to mechanical loads in an isoform-dependent manner, resulting in their ability to dynamically adapt to a range of motile challenges. Here, we reveal the ...Myosins adjust their power outputs in response to mechanical loads in an isoform-dependent manner, resulting in their ability to dynamically adapt to a range of motile challenges. Here, we reveal the structural basis for force-sensing based on near-atomic resolution structures of one rigor and two ADP-bound states of myosin-IB (myo1b) bound to actin, determined by cryo-electron microscopy. The two ADP-bound states are separated by a 25° rotation of the lever. The lever of the first ADP state is rotated toward the pointed end of the actin filament and forms a previously unidentified interface with the N-terminal subdomain, which constitutes the upper half of the nucleotide-binding cleft. This pointed-end orientation of the lever blocks ADP release by preventing the N-terminal subdomain from the pivoting required to open the nucleotide binding site, thus revealing how myo1b is inhibited by mechanical loads that restrain lever rotation. The lever of the second ADP state adopts a rigor-like orientation, stabilized by class-specific elements of myo1b. We identify a role for this conformation as an intermediate in the ADP release pathway. Moreover, comparison of our structures with other myosins reveals structural diversity in the actomyosin binding site, and we reveal the high-resolution structure of actin-bound phalloidin, a potent stabilizer of filamentous actin. These results provide a framework to understand the spectrum of force-sensing capacities among the myosin superfamily. #1: Journal: Proc. Natl. Acad. Sci. U.S.A. / Year: 2014Title: A vertebrate myosin-I structure reveals unique insights into myosin mechanochemical tuning. Authors: Shuman, H. / Greenberg, M.J. / Zwolak, A. / Lin, T. / Sindelar, C.V. / Dominguez, R. / Ostap, E.M. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5v7x.cif.gz | 183.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5v7x.ent.gz | 144.9 KB | Display | PDB format |
| PDBx/mmJSON format | 5v7x.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/v7/5v7xftp://data.pdbj.org/pub/pdb/validation_reports/v7/5v7x | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  7329C  7330C  7331C  6c1dC  6c1gC  6c1hC C: citing same article ( |
|---|---|
| Similar structure data |







-Links
 PDBj
PDBj























- Assembly
Assembly
| Deposited unit | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 86954.969 Da / Num. of mol.: 1 / Fragment: UNP residues 1-728 Source method: isolated from a genetically manipulated source Source: (gene. exp.)   Spodoptera frugiperda (fall armyworm) / References: UniProt: Q05096 Spodoptera frugiperda (fall armyworm) / References: UniProt: Q05096 |
|---|---|
| #2: Protein | Mass: 16852.545 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: CALM1, CALM, CAM, CAM1 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: P0DP23 |
| #3: Chemical | ChemComp-SO4 / 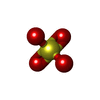  Mass: 96.063 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: SO4 Mass: 96.063 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: SO4 |
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 3.05 Å3/Da / Density % sol: 59.7 % |
|---|---|
| Crystal grow | Temperature: 277 K / Method: vapor diffusion, hanging drop Details: 10% PEG 3350, 100 mM LiOOCCH3, 100 mM HEPES, pH 7.0, 20 mM Mg2SO4 |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: NSLS / Beamline: X25 / Wavelength: 1 Å |
| Detector | Type: ADSC QUANTUM 315 / Detector: CCD / Date: Nov 7, 2012 Details: Monochromator: Double silicon(111) crystal monochromator with cryogenically-cooled first crystal and sagittally-bent second crystal horizontally-focusing at 3.3:1 demagnification. Mirror: ...Details: Monochromator: Double silicon(111) crystal monochromator with cryogenically-cooled first crystal and sagittally-bent second crystal horizontally-focusing at 3.3:1 demagnification. Mirror: Meridionally-bent fused silica mirror with palladium and uncoated stripes vertically-focusing at 6.6:1 demagnification. |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1 Å / Relative weight: 1 |
| Reflection twin | Operator: h,-k,-l / Fraction: 0.39 |
| Reflection | Resolution: 2.799→39.504 Å / Num. obs: 30781 / % possible obs: 95.8 % / Redundancy: 7.3 % / Net I/σ(I): 9.5 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Resolution: 3.141→39.504 Å / Cross valid method: THROUGHOUT / σ(F): 37.84 / Phase error: 26.56 / Stereochemistry target values: TWIN_LSQ_F
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 3.141→39.504 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
