 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5u8s | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Structure of eukaryotic CMG helicase at a replication fork | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | REPLICATION / CMG helicase / replisome / origin initiation / DNA polymerase / DNA replication | |||||||||
| Function / homology |  Function and homology information Function and homology informationUnwinding of DNA / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / DNA strand elongation involved in mitotic DNA replication / GINS complex / MCM complex binding / mitotic DNA replication preinitiation complex assembly / nuclear DNA replication / premeiotic DNA replication ...Unwinding of DNA / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / DNA strand elongation involved in mitotic DNA replication / GINS complex / MCM complex binding / mitotic DNA replication preinitiation complex assembly / nuclear DNA replication / premeiotic DNA replication / replication fork protection complex / pre-replicative complex assembly involved in nuclear cell cycle DNA replication / Activation of the pre-replicative complex / mitotic DNA replication / CMG complex / nuclear pre-replicative complex / Activation of ATR in response to replication stress / DNA replication preinitiation complex / MCM complex / double-strand break repair via break-induced replication / mitotic DNA replication initiation / single-stranded DNA helicase activity / silent mating-type cassette heterochromatin formation / regulation of DNA-templated DNA replication initiation / DNA strand elongation involved in DNA replication / nuclear replication fork / DNA replication origin binding / DNA replication initiation / subtelomeric heterochromatin formation / DNA helicase activity / transcription elongation by RNA polymerase II / helicase activity / DNA-templated DNA replication / heterochromatin formation / single-stranded DNA binding / DNA helicase / chromosome, telomeric region / DNA replication / DNA damage response / chromatin binding / ATP hydrolysis activity / zinc ion binding / nucleoplasm / ATP binding / nucleus / cytoplasm Similarity search - Function | |||||||||
| Biological species |  synthetic construct (others) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 6.1 Å | |||||||||
 Authors Authors | Li, H. / Li, B. / Georgescu, R. / Yuan, Z. / Santos, R. / Sun, J. / Zhang, D. / Yurieva, O. / O'Donnell, M.E. | |||||||||
| Funding support |  United States, 2items United States, 2items
| |||||||||
 Citation Citation | Journal: Proc Natl Acad Sci U S A / Year: 2017 Title: Structure of eukaryotic CMG helicase at a replication fork and implications to replisome architecture and origin initiation. Authors: Roxana Georgescu / Zuanning Yuan / Lin Bai / Ruda de Luna Almeida Santos / Jingchuan Sun / Dan Zhang / Olga Yurieva / Huilin Li / Michael E O'Donnell / Abstract: The eukaryotic CMG (Cdc45, Mcm2-7, GINS) helicase consists of the Mcm2-7 hexameric ring along with five accessory factors. The Mcm2-7 heterohexamer, like other hexameric helicases, is shaped like a ...The eukaryotic CMG (Cdc45, Mcm2-7, GINS) helicase consists of the Mcm2-7 hexameric ring along with five accessory factors. The Mcm2-7 heterohexamer, like other hexameric helicases, is shaped like a ring with two tiers, an N-tier ring composed of the N-terminal domains, and a C-tier of C-terminal domains; the C-tier contains the motor. In principle, either tier could translocate ahead of the other during movement on DNA. We have used cryo-EM single-particle 3D reconstruction to solve the structure of CMG in complex with a DNA fork. The duplex stem penetrates into the central channel of the N-tier and the unwound leading single-strand DNA traverses the channel through the N-tier into the C-tier motor, 5'-3' through CMG. Therefore, the N-tier ring is pushed ahead by the C-tier ring during CMG translocation, opposite the currently accepted polarity. The polarity of the N-tier ahead of the C-tier places the leading Pol ε below CMG and Pol α-primase at the top of CMG at the replication fork. Surprisingly, the new N-tier to C-tier polarity of translocation reveals an unforeseen quality-control mechanism at the origin. Thus, upon assembly of head-to-head CMGs that encircle double-stranded DNA at the origin, the two CMGs must pass one another to leave the origin and both must remodel onto opposite strands of single-stranded DNA to do so. We propose that head-to-head motors may generate energy that underlies initial melting at the origin. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5u8s.cif.gz | 1 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5u8s.ent.gz | 834.8 KB | Display | PDB format |
| PDBx/mmJSON format | 5u8s.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 5u8s_validation.pdf.gz | 1.3 MB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 5u8s_full_validation.pdf.gz | 1.8 MB | Display | |
| Data in XML | 5u8s_validation.xml.gz | 226.5 KB | Display | |
| Data in CIF | 5u8s_validation.cif.gz | 324.8 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/u8/5u8sftp://data.pdbj.org/pub/pdb/validation_reports/u8/5u8s | HTTPS FTP |
-Related structure data
| Related structure data |  8518MC  8519C  8520C  5u8tC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |


-Links
 PDBj
PDBj






































- Assembly
Assembly
| Deposited unit | 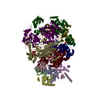
|
|---|---|
| 1 |
|
-Components
-DNA replication complex GINS protein ... , 4 types, 4 molecules ABCD
| #1: Protein | Mass: 24230.576 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: PSF1, YDR013W, PZA208, YD8119.18 / Production host: |
|---|---|
| #2: Protein | Mass: 25096.807 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: PSF2, YJL072C, HRF213, J1086 / Production host: |
| #3: Protein | Mass: 21977.135 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: PSF3, YOL146W / Production host: |
| #4: Protein | Mass: 33983.617 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: SLD5, YDR489W / Production host: |
-Protein , 2 types, 2 molecules E5
| #5: Protein | Mass: 74324.836 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: CDC45, SLD4, YLR103C, L8004.11 / Production host: |
|---|---|
| #11: Protein | Mass: 86505.734 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: MCM5, CDC46, YLR274W, L9328.1 / Production host: |
-DNA chain , 2 types, 2 molecules FG
| #6: DNA chain | Mass: 7930.117 Da / Num. of mol.: 1 / Source method: obtained synthetically / Details: fork DNA_a (26-MER) / Source: (synth.) synthetic construct (others) |
|---|---|
| #7: DNA chain | Mass: 4279.804 Da / Num. of mol.: 1 / Source method: obtained synthetically / Details: fork DNA-b (14-mer) / Source: (synth.) synthetic construct (others) |
-DNA replication licensing factor ... , 5 types, 5 molecules 23467
| #8: Protein | Mass: 98911.539 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: MCM2, YBL023C, YBL0438 / Production host: |
|---|---|
| #9: Protein | Mass: 107653.508 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: MCM3, YEL032W, SYGP-ORF23 / Production host: |
| #10: Protein | Mass: 105096.273 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: MCM4, CDC54, HCD21, YPR019W, YP9531.13 / Production host: |
| #12: Protein | Mass: 113110.211 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: MCM6, YGL201C / Production host: |
| #13: Protein | Mass: 95049.875 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: MCM7, CDC47, YBR202W, YBR1441 / Production host: |
-Non-polymers , 1 types, 3 molecules 
| #14: Chemical |  Mass: 507.181 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM |
|---|
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: CMG-biotinylated fork DNA-Streptavidin / Type: COMPLEX / Entity ID: #1-#13 / Source: RECOMBINANT |
|---|---|
| Molecular weight | Value: 0.75 MDa / Experimental value: NO |
| Source (natural) | Organism: |
| Source (recombinant) | Organism: |
| Buffer solution | pH: 7.5 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD |
| Image recording | Electron dose: 10 e/Å2 / Detector mode: SUPER-RESOLUTION / Film or detector model: GATAN K2 SUMMIT (4k x 4k) |
- Processing
Processing
| Software | Name: PHENIX / Version: 1.9_1692 / Classification: refinement | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 6.1 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 243796 / Symmetry type: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: RIGID BODY FIT / Space: REAL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Resolution: 6.1→6.1 Å / SU ML: 1.98 / Phase error: 57.46 / Stereochemistry target values: MLHL
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
