 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5nl0 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Crystal structure of a 197-bp palindromic 601L nucleosome in complex with linker histone H1 | |||||||||||||||||||||
 要素 要素 |
| |||||||||||||||||||||
 キーワード キーワード | CHROMATIN BINDING PROTEIN / DNA / nucleosome / chromatin / linker histones / histone H1 / CHROMATIN BINDING PROTEIN - DNA complex | |||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報negative regulation of DNA recombination / chromosome condensation / nucleosomal DNA binding / structural constituent of chromatin / nucleosome / nucleosome assembly / double-stranded DNA binding / protein heterodimerization activity / DNA binding / nucleoplasm / nucleus 類似検索 - 分子機能 | |||||||||||||||||||||
| 生物種 | synthetic construct (人工物) | |||||||||||||||||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 5.4 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 5.4 Å | |||||||||||||||||||||
 データ登録者 データ登録者 | Garcia-Saez, I. / Petosa, C. / Dimitrov, S. | |||||||||||||||||||||
| 資金援助 |  フランス, 6件 フランス, 6件
| |||||||||||||||||||||
 引用 引用 | ジャーナル: Mol Cell / 年: 2017 タイトル: Structure and Dynamics of a 197 bp Nucleosome in Complex with Linker Histone H1. 著者: Jan Bednar / Isabel Garcia-Saez / Ramachandran Boopathi / Amber R Cutter / Gabor Papai / Anna Reymer / Sajad H Syed / Imtiaz Nisar Lone / Ognyan Tonchev / Corinne Crucifix / Hervé Menoni / ...著者: Jan Bednar / Isabel Garcia-Saez / Ramachandran Boopathi / Amber R Cutter / Gabor Papai / Anna Reymer / Sajad H Syed / Imtiaz Nisar Lone / Ognyan Tonchev / Corinne Crucifix / Hervé Menoni / Christophe Papin / Dimitrios A Skoufias / Hitoshi Kurumizaka / Richard Lavery / Ali Hamiche / Jeffrey J Hayes / Patrick Schultz / Dimitar Angelov / Carlo Petosa / Stefan Dimitrov /   要旨: Linker histones associate with nucleosomes to promote the formation of higher-order chromatin structure, but the underlying molecular details are unclear. We investigated the structure of a 197 bp ...Linker histones associate with nucleosomes to promote the formation of higher-order chromatin structure, but the underlying molecular details are unclear. We investigated the structure of a 197 bp nucleosome bearing symmetric 25 bp linker DNA arms in complex with vertebrate linker histone H1. We determined electron cryo-microscopy (cryo-EM) and crystal structures of unbound and H1-bound nucleosomes and validated these structures by site-directed protein cross-linking and hydroxyl radical footprinting experiments. Histone H1 shifts the conformational landscape of the nucleosome by drawing the two linkers together and reducing their flexibility. The H1 C-terminal domain (CTD) localizes primarily to a single linker, while the H1 globular domain contacts the nucleosome dyad and both linkers, associating more closely with the CTD-distal linker. These findings reveal that H1 imparts a strong degree of asymmetry to the nucleosome, which is likely to influence the assembly and architecture of higher-order structures. | |||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5nl0.cif.gz | 1.1 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5nl0.ent.gz | 935.7 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5nl0.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5nl0_validation.pdf.gz | 497.9 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5nl0_full_validation.pdf.gz | 528 KB | 表示 | |
| XML形式データ | 5nl0_validation.xml.gz | 32.9 KB | 表示 | |
| CIF形式データ | 5nl0_validation.cif.gz | 53.4 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/nl/5nl0ftp://data.pdbj.org/pub/pdb/validation_reports/nl/5nl0 | HTTPS FTP |
-関連構造データ
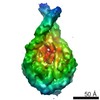






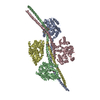
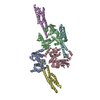

-リンク
 PDBj
PDBj






































- 集合体
集合体
| 登録構造単位 | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 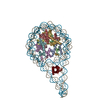
| ||||||||
| 2 | 
| ||||||||
| 単位格子 |
|
-要素
-タンパク質 , 5種, 13分子 AEKBFLCGMDHNZ
| #1: タンパク質 | 分子量: 15303.930 Da / 分子数: 3 / 由来タイプ: 組換発現 詳細: The core histones in the uploaded coordinates are those from X. laevis while those in the crystal are human. The reason for the discrepancy is that the structure was solved using pdb 3UT9 as ...詳細: The core histones in the uploaded coordinates are those from X. laevis while those in the crystal are human. The reason for the discrepancy is that the structure was solved using pdb 3UT9 as a molecular replacement template (because it had the same 601L DNA sequence and therefore was the closest high-res structure available). 由来: (組換発現) 発現宿主:  #2: タンパク質 | 分子量: 11263.231 Da / 分子数: 3 / 由来タイプ: 組換発現 詳細: The core histones in the uploaded coordinates are those from X. laevis while those in the crystal are human. The reason for the discrepancy is that the structure was solved using pdb 3UT9 as ...詳細: The core histones in the uploaded coordinates are those from X. laevis while those in the crystal are human. The reason for the discrepancy is that the structure was solved using pdb 3UT9 as a molecular replacement template (because it had the same 601L DNA sequence and therefore was the closest high-res structure available). 由来: (組換発現) 発現宿主: #3: タンパク質 | 分子量: 13978.241 Da / 分子数: 3 / 由来タイプ: 組換発現 詳細: The core histones in the uploaded coordinates are those from X. laevis while those in the crystal are human. The reason for the discrepancy is that the structure was solved using pdb 3UT9 as ...詳細: The core histones in the uploaded coordinates are those from X. laevis while those in the crystal are human. The reason for the discrepancy is that the structure was solved using pdb 3UT9 as a molecular replacement template (because it had the same 601L DNA sequence and therefore was the closest high-res structure available). 由来: (組換発現) 発現宿主: #4: タンパク質 | 分子量: 13524.752 Da / 分子数: 3 / 由来タイプ: 組換発現 詳細: The core histones in the uploaded coordinates are those from X. laevis while those in the crystal are human. The reason for the discrepancy is that the structure was solved using pdb 3UT9 as ...詳細: The core histones in the uploaded coordinates are those from X. laevis while those in the crystal are human. The reason for the discrepancy is that the structure was solved using pdb 3UT9 as a molecular replacement template (because it had the same 601L DNA sequence and therefore was the closest high-res structure available). 由来: (組換発現) 発現宿主: #7: タンパク質 | | 分子量: 21129.502 Da / 分子数: 1 / 由来タイプ: 組換発現 詳細: Only the H1.0b globular domain was visible in the structure. The coordinates of this chain are those of a homology model built from chicken GH5 (pdb 1HST). 由来: (組換発現) 遺伝子: h1f0-b / 発現宿主: |
|---|
-DNA鎖 , 2種, 4分子 ISJT
| #5: DNA鎖 | 分子量: 60824.777 Da / 分子数: 2 / 由来タイプ: 組換発現 詳細: There are only 193 bp in the structure whereas there are 197 bp in the crystal; putatively this could be because two base pairs at the tip of each linker are denatured and therefore not visible. 由来: (組換発現) synthetic construct (人工物) / 発現宿主: #6: DNA鎖 | 分子量: 60815.762 Da / 分子数: 2 / 由来タイプ: 組換発現 詳細: There are only 193 bp in the structure whereas there are 197 bp in the crystal; putatively this could be because two base pairs at the tip of each linker are denatured and therefore not visible. 由来: (組換発現) synthetic construct (人工物) / 発現宿主: |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 3.21 Å3/Da / 溶媒含有率: 61.68 % / 解説: Long thin rods. |
|---|---|
| 結晶化 | 温度: 293.15 K / 手法: 蒸気拡散法, ハンギングドロップ法 / pH: 6.4 詳細: Mix of equal volumes of the nucleosome/H1 complex (25-30 microM) and a crystallization solution composed of MPD (6% v/v), 50 mM NaCl, and 50 mM sodium potassium phosphate pH 6.4. |
-データ収集
| 回折 | 平均測定温度: 100 K |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: ESRF / ビームライン: ID29 / 波長: 0.99987 Å |
| 検出器 | タイプ: DECTRIS PILATUS 6M / 検出器: PIXEL / 日付: 2015年8月29日 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 0.99987 Å / 相対比: 1 |
| 反射 | 解像度: 5.4→49.13 Å / Num. obs: 15641 / % possible obs: 99.5 % / 冗長度: 4.5 % / CC1/2: 0.997 / Rmerge(I) obs: 0.091 / Rpim(I) all: 0.072 / Net I/σ(I): 9.1 |
| 反射 シェル | 解像度: 5.4→6.04 Å / 冗長度: 4.4 % / Rmerge(I) obs: 0.685 / Mean I/σ(I) obs: 2.1 / Num. unique obs: 4370 / CC1/2: 0.686 / Rpim(I) all: 0.553 / % possible all: 99.6 |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 開始モデル: 3UT9 解像度: 5.4→49.128 Å / SU ML: 0.85 / 交差検証法: FREE R-VALUE / σ(F): 1.35 / 位相誤差: 31.59 詳細: The asymmetric unit contains a complete H1/nucleosome complex (molecule A) and half of a second complex (molecule B). Rigid-body and TLS refinement in Phenix were performed. Linker DNA was ...詳細: The asymmetric unit contains a complete H1/nucleosome complex (molecule A) and half of a second complex (molecule B). Rigid-body and TLS refinement in Phenix were performed. Linker DNA was extended from the nucleosome core as B-form DNA, adjusted into density using Coot and subjected to rigid body and TLS refinement in Phenix, resulting in improved R-values. The 2Fo-Fc and Fo-Fc maps calculated using phases from the NCP and linker DNA revealed additional density corresponding to the GH1 domain in both complexes A and B. The structure of X. laevis GH1.0b was modeled by homology with the crystal structure of chicken GH5 (PDB 1HST), which shares 80% amino acid sequence identity with X. laevis GH1.0b.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 5.4→49.128 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル |
|
