 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-5m50: Mechanism of microtubule minus-end recognition and protection by ... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5m50 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Mechanism of microtubule minus-end recognition and protection by CAMSAP proteins | |||||||||||||||||||||
 Components Components |
| |||||||||||||||||||||
 Keywords Keywords | STRUCTURAL PROTEIN / CAMSAP CKK Microtubule Tubulin | |||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationregulation of organelle organization / zonula adherens maintenance / microtubule minus-end / Intraflagellar transport / microtubule anchoring / Resolution of Sister Chromatid Cohesion / EML4 and NUDC in mitotic spindle formation / regulation of Golgi organization / COPI-independent Golgi-to-ER retrograde traffic / COPI-mediated anterograde transport ...regulation of organelle organization / zonula adherens maintenance / microtubule minus-end / Intraflagellar transport / microtubule anchoring / Resolution of Sister Chromatid Cohesion / EML4 and NUDC in mitotic spindle formation / regulation of Golgi organization / COPI-independent Golgi-to-ER retrograde traffic / COPI-mediated anterograde transport / protein transport along microtubule / microtubule minus-end binding / establishment or maintenance of microtubule cytoskeleton polarity / MHC class II antigen presentation / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / cilium movement / Aggrephagy / epithelial cell-cell adhesion / zonula adherens / Separation of Sister Chromatids / Loss of Nlp from mitotic centrosomes / Recruitment of mitotic centrosome proteins and complexes / Loss of proteins required for interphase microtubule organization from the centrosome / Anchoring of the basal body to the plasma membrane / AURKA Activation by TPX2 / Recruitment of NuMA to mitotic centrosomes / Regulation of PLK1 Activity at G2/M Transition / negative regulation of microtubule depolymerization / establishment of epithelial cell apical/basal polarity / embryo development ending in birth or egg hatching / motile cilium / positive regulation of axon guidance / regulation of focal adhesion assembly / spectrin binding / regulation of microtubule polymerization / axoneme / cellular response to interleukin-4 / microtubule-based process / cytoplasmic microtubule / cytoplasmic microtubule organization / regulation of cell migration / regulation of microtubule cytoskeleton organization / neuron migration / microtubule cytoskeleton organization / structural constituent of cytoskeleton / neuron projection development / neuron differentiation / actin filament binding / in utero embryonic development / microtubule cytoskeleton / mitotic cell cycle / double-stranded RNA binding / cilium / microtubule / Hydrolases; Acting on acid anhydrides; Acting on GTP to facilitate cellular and subcellular movement / ciliary basal body / calmodulin binding / protein heterodimerization activity / hydrolase activity / GTPase activity / centrosome / ubiquitin protein ligase binding / GTP binding / metal ion binding / cytosol / cytoplasm Similarity search - Function | |||||||||||||||||||||
| Biological species |  | |||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 5.3 Å | |||||||||||||||||||||
 Authors Authors | Akhmanova, A. / Moores, C.A. / Baldus, M. / Steinmetz, M.O. / Topf, M. / Roberts, A.J. / Grant, B.J. / Scarabelli, G. / Joseph, A.-P. / van Hooff, J.J.E. ...Akhmanova, A. / Moores, C.A. / Baldus, M. / Steinmetz, M.O. / Topf, M. / Roberts, A.J. / Grant, B.J. / Scarabelli, G. / Joseph, A.-P. / van Hooff, J.J.E. / Houben, K. / Hua, S. / Luo, Y. / Stangier, M.M. / Jiang, K. / Atherton, J. | |||||||||||||||||||||
| Funding support |  United Kingdom, United Kingdom,  United States, United States,  Netherlands, Netherlands,  Switzerland, 6items Switzerland, 6items
| |||||||||||||||||||||
 Citation Citation | Journal: Nat Struct Mol Biol / Year: 2017 Title: A structural model for microtubule minus-end recognition and protection by CAMSAP proteins. Authors: Joseph Atherton / Kai Jiang / Marcel M Stangier / Yanzhang Luo / Shasha Hua / Klaartje Houben / Jolien J E van Hooff / Agnel-Praveen Joseph / Guido Scarabelli / Barry J Grant / Anthony J ...Authors: Joseph Atherton / Kai Jiang / Marcel M Stangier / Yanzhang Luo / Shasha Hua / Klaartje Houben / Jolien J E van Hooff / Agnel-Praveen Joseph / Guido Scarabelli / Barry J Grant / Anthony J Roberts / Maya Topf / Michel O Steinmetz / Marc Baldus / Carolyn A Moores / Anna Akhmanova / Abstract: CAMSAP and Patronin family members regulate microtubule minus-end stability and localization and thus organize noncentrosomal microtubule networks, which are essential for cell division, polarization ...CAMSAP and Patronin family members regulate microtubule minus-end stability and localization and thus organize noncentrosomal microtubule networks, which are essential for cell division, polarization and differentiation. Here, we found that the CAMSAP C-terminal CKK domain is widely present among eukaryotes and autonomously recognizes microtubule minus ends. Through a combination of structural approaches, we uncovered how mammalian CKK binds between two tubulin dimers at the interprotofilament interface on the outer microtubule surface. In vitro reconstitution assays combined with high-resolution fluorescence microscopy and cryo-electron tomography suggested that CKK preferentially associates with the transition zone between curved protofilaments and the regular microtubule lattice. We propose that minus-end-specific features of the interprotofilament interface at this site serve as the basis for CKK's minus-end preference. The steric clash between microtubule-bound CKK and kinesin motors explains how CKK protects microtubule minus ends against kinesin-13-induced depolymerization and thus controls the stability of free microtubule minus ends. | |||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5m50.cif.gz | 335.8 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5m50.ent.gz | 267.6 KB | Display | PDB format |
| PDBx/mmJSON format | 5m50.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/m5/5m50ftp://data.pdbj.org/pub/pdb/validation_reports/m5/5m50 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  4154MC  3444C  4156C  5lznC  5m54C  5m5cC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj



- Assembly
Assembly
| Deposited unit | 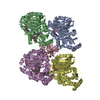
|
|---|---|
| 1 |
|
-Components
-Protein , 3 types, 5 molecules DAEBC
| #1: Protein | Mass: 48769.988 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #2: Protein | Mass: 47940.945 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #3: Protein | | Mass: 13431.391 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.)  |
|---|
-Non-polymers , 5 types, 16 molecules 


| #4: Chemical |  Mass: 523.180 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM Mass: 523.180 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM#5: Chemical |  Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg#6: Chemical |  Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM#7: Chemical |  Mass: 853.906 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C47H51NO14 / Comment: medication, chemotherapy*YM Mass: 853.906 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C47H51NO14 / Comment: medication, chemotherapy*YM#8: Water | ChemComp-HOH / | Mass: 18.015 Da / Num. of mol.: 8 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: FILAMENT / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source (natural) |
| ||||||||||||||||||||||||
| Source (recombinant) | Organism: | ||||||||||||||||||||||||
| Buffer solution | pH: 6.8 / Details: BRB80 | ||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES / Details: 13pf Microtubules | ||||||||||||||||||||||||
| Specimen support | Grid material: COPPER / Grid type: C-flat-2/2 | ||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK III / Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Tecnai Polara / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI POLARA 300 |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD |
| Image recording | Electron dose: 25 e/Å2 / Detector mode: INTEGRATING / Film or detector model: DIRECT ELECTRON DE-20 (5k x 3k) / Details: 25e-/A2 used in final reconstuctions |
- Processing
Processing
| Software | Name: PHENIX / Version: 1.11.1_2575: / Classification: refinement | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 5.3 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 6530 Details: Gold-standard noise substitution test used to asses for over fitting (Chen et al., 2013) Symmetry type: POINT | ||||||||||||||||||||||||
| Atomic model building | Protocol: OTHER | ||||||||||||||||||||||||
| Refine LS restraints |
|
