 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5hyn | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Structure of Human Polycomb Repressive Complex 2 (PRC2) with oncogenic histone H3K27M peptide | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | TRANSFERASE / chromatin modification complex | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報protein localization to pericentric heterochromatin / hepatocyte homeostasis / cellular response to trichostatin A / regulation of gliogenesis / negative regulation of striated muscle cell differentiation / regulation of kidney development / [histone H3]-lysine27 N-trimethyltransferase / response to tetrachloromethane / negative regulation of keratinocyte differentiation / histone H3K27 trimethyltransferase activity ...protein localization to pericentric heterochromatin / hepatocyte homeostasis / cellular response to trichostatin A / regulation of gliogenesis / negative regulation of striated muscle cell differentiation / regulation of kidney development / [histone H3]-lysine27 N-trimethyltransferase / response to tetrachloromethane / negative regulation of keratinocyte differentiation / histone H3K27 trimethyltransferase activity / negative regulation of retinoic acid receptor signaling pathway / cerebellar cortex development / primary miRNA binding / random inactivation of X chromosome / regulation of adaxial/abaxial pattern formation / skeletal muscle satellite cell maintenance involved in skeletal muscle regeneration / regulatory ncRNA-mediated heterochromatin formation / negative regulation of cardiac muscle cell proliferation / histone H3K27 methyltransferase activity / sex chromatin / ubiquitin-modified histone reader activity / positive regulation of cell cycle G1/S phase transition / facultative heterochromatin formation / genomic imprinting / ESC/E(Z) complex / negative regulation of stem cell differentiation / RSC-type complex / protein-lysine N-methyltransferase activity / cardiac muscle hypertrophy in response to stress / chromatin silencing complex / histone H3K9me2/3 reader activity / pronucleus / positive regulation of dendrite development / G1 to G0 transition / histone H3 methyltransferase activity / histone methyltransferase activity / DNA methylation-dependent constitutive heterochromatin formation / negative regulation of G1/S transition of mitotic cell cycle / spinal cord development / lncRNA binding / negative regulation of gene expression, epigenetic / synaptic transmission, GABAergic / Transcriptional Regulation by E2F6 / : / positive regulation of MAP kinase activity / histone methyltransferase complex / oligodendrocyte differentiation / negative regulation of transcription elongation by RNA polymerase II / positive regulation of GTPase activity / positive regulation of protein serine/threonine kinase activity / negative regulation of cell differentiation / cardiac muscle cell proliferation / positive regulation of epithelial to mesenchymal transition / ribonucleoprotein complex binding / subtelomeric heterochromatin formation / RNA polymerase II core promoter sequence-specific DNA binding / pericentric heterochromatin / nucleosome binding / keratinocyte differentiation / Chromatin modifying enzymes / spleen development / protein localization to chromatin / liver regeneration / negative regulation of cytokine production involved in inflammatory response / telomere organization / Interleukin-7 signaling / RNA Polymerase I Promoter Opening / Assembly of the ORC complex at the origin of replication / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / SUMOylation of chromatin organization proteins / thymus development / cellular response to leukemia inhibitory factor / DNA methylation / central nervous system development / B cell differentiation / epigenetic regulation of gene expression / Condensation of Prophase Chromosomes / ubiquitin binding / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / Regulation of PTEN gene transcription / transcription corepressor binding / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / hippocampus development / stem cell differentiation / HDACs deacetylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / enzyme activator activity / promoter-specific chromatin binding / RNA Polymerase I Promoter Escape / liver development / Transcriptional regulation by small RNAs / G1/S transition of mitotic cell cycle / Formation of the beta-catenin:TCF transactivating complex / protein-DNA complex / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 |  X線回折 / シンクロトロン / 単波長異常分散 / 解像度: 2.95 Å X線回折 / シンクロトロン / 単波長異常分散 / 解像度: 2.95 Å | ||||||
 データ登録者 データ登録者 | Zhang, Y. / Justin, N. / Wilson, J.R. / Gamblin, S.J. | ||||||
| 資金援助 |  英国, 1件 英国, 1件
| ||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2016 タイトル: Structural basis of oncogenic histone H3K27M inhibition of human polycomb repressive complex 2. 著者: Justin, N. / Zhang, Y. / Tarricone, C. / Martin, S.R. / Chen, S. / Underwood, E. / De Marco, V. / Haire, L.F. / Walker, P.A. / Reinberg, D. / Wilson, J.R. / Gamblin, S.J. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5hyn.cif.gz | 1.7 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5hyn.ent.gz | 1.4 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5hyn.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5hyn_validation.pdf.gz | 2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5hyn_full_validation.pdf.gz | 2.1 MB | 表示 | |
| XML形式データ | 5hyn_validation.xml.gz | 184.5 KB | 表示 | |
| CIF形式データ | 5hyn_validation.cif.gz | 230.8 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/hy/5hynftp://data.pdbj.org/pub/pdb/validation_reports/hy/5hyn | HTTPS FTP |
-関連構造データ






-リンク
 PDBj
PDBj















- 集合体
集合体
| 登録構造単位 | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| 3 | 
| ||||||||
| 4 | 
| ||||||||
| 5 | 
| ||||||||
| 6 | 
| ||||||||
| 7 | 
| ||||||||
| 8 | 
| ||||||||
| 単位格子 |
|
-要素
-タンパク質 , 1種, 4分子 AFKQ
| #1: タンパク質 | 分子量: 85492.297 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: EZH2, KMT6 / 発現宿主:  |
|---|
-Polycomb protein ... , 2種, 8分子 BGLRCHMS
| #2: タンパク質 | 分子量: 42314.145 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: EED / 発現宿主: #3: タンパク質 | 分子量: 15249.510 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: SUZ12, CHET9, JJAZ1, KIAA0160 / 発現宿主: |
|---|
-タンパク質・ペプチド , 2種, 8分子 DIOTEJPU
| #4: タンパク質・ペプチド | 分子量: 1234.426 Da / 分子数: 4 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) / 参照: UniProt: P68431*PLUS#5: タンパク質・ペプチド | 分子量: 1505.743 Da / 分子数: 4 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) / 参照: UniProt: Q92833*PLUS |
|---|
-非ポリマー , 2種, 36分子 
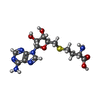
| #6: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 32 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 32 / 由来タイプ: 合成 / 式: Zn#7: 化合物 | ChemComp-SAH /  分子量: 384.411 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C14H20N6O5S 分子量: 384.411 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C14H20N6O5S |
|---|
-詳細
| Has protein modification | Y |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 2.82 Å3/Da / 溶媒含有率: 56.33 % |
|---|---|
| 結晶化 | 温度: 291 K / 手法: 蒸気拡散法, シッティングドロップ法 / pH: 6.5 / 詳細: 25% PEG3350, 400mM Ammonian Citrate pH6.5 / PH範囲: 6.5-6.8 |
-データ収集
| 回折 | 平均測定温度: 100 K |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: Diamond / ビームライン: I03 / 波長: 0.9763 Å |
| 検出器 | タイプ: DECTRIS PILATUS3 6M / 検出器: PIXEL / 日付: 2015年10月8日 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 0.9763 Å / 相対比: 1 |
| 反射 | 解像度: 2.94→104.43 Å / Num. obs: 132394 / % possible obs: 99.9 % / 冗長度: 5.6 % / CC1/2: 0.996 / Rmerge(I) obs: 0.165 / Net I/σ(I): 9.9 |
| 反射 シェル | 解像度: 2.94→3.02 Å / 冗長度: 5.6 % / Rmerge(I) obs: 1.607 / Mean I/σ(I) obs: 1.4 / % possible all: 99.9 |
- 解析
解析
| ソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 単波長異常分散 / 解像度: 2.95→95.013 Å / SU ML: 0.5 / 交差検証法: FREE R-VALUE / σ(F): 1.33 / 位相誤差: 32.32
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 2.95→95.013 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ |
|