 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 4ujd | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | mammalian 80S HCV-IRES initiation complex with eIF5B PRE-like state | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | RIBOSOME / TRANSLATION INITIATION / HEPATITIS C VIRUS INTERNAL RIBOSOME ENTRY SITE / EUKARYOTIC INITIATION FACTOR 5B | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報protein-synthesizing GTPase / translation initiation factor activity / GTPase activity / GTP binding / mitochondrion / metal ion binding 類似検索 - 分子機能 | ||||||||||||
| 生物種 |   HEPATITIS C VIRUS (ウイルス) HEPATITIS C VIRUS (ウイルス) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 8.9 Å | ||||||||||||
 データ登録者 データ登録者 | Yamamoto, H. / Unbehaun, A. / Loerke, J. / Behrmann, E. / Marianne, C. / Burger, J. / Mielke, T. / Spahn, C.M.T. | ||||||||||||
 引用 引用 | ジャーナル: Nat Struct Mol Biol / 年: 2014 タイトル: Structure of the mammalian 80S initiation complex with initiation factor 5B on HCV-IRES RNA. 著者: Hiroshi Yamamoto / Anett Unbehaun / Justus Loerke / Elmar Behrmann / Marianne Collier / Jörg Bürger / Thorsten Mielke / Christian M T Spahn /  要旨: The universally conserved eukaryotic initiation factor (eIF) 5B, a translational GTPase, is essential for canonical translation initiation. It is also required for initiation facilitated by the ...The universally conserved eukaryotic initiation factor (eIF) 5B, a translational GTPase, is essential for canonical translation initiation. It is also required for initiation facilitated by the internal ribosomal entry site (IRES) of hepatitis C virus (HCV) RNA. eIF5B promotes joining of 60S ribosomal subunits to 40S ribosomal subunits bound by initiator tRNA (Met-tRNAi(Met)). However, the exact molecular mechanism by which eIF5B acts has not been established. Here we present cryo-EM reconstructions of the mammalian 80S-HCV-IRES-Met-tRNAi(Met)-eIF5B-GMPPNP complex. We obtained two substates distinguished by the rotational state of the ribosomal subunits and the configuration of initiator tRNA in the peptidyl (P) site. Accordingly, a combination of conformational changes in the 80S ribosome and in initiator tRNA facilitates binding of the Met-tRNAi(Met) to the 60S P site and redefines the role of eIF5B as a tRNA-reorientation factor. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 4ujd.cif.gz | 4.8 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb4ujd.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 4ujd.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/uj/4ujdftp://data.pdbj.org/pub/pdb/validation_reports/uj/4ujd | HTTPS FTP |
|---|
-関連構造データ
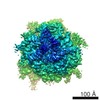





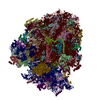
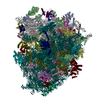
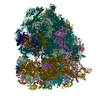


-リンク
 PDBj
PDBj






























- 集合体
集合体
| 登録構造単位 | 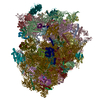
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 6種, 6分子 A2A3A4BABCC1
| #1: RNA鎖 | 分子量: 1625917.625 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #2: RNA鎖 | 分子量: 62616.344 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #3: RNA鎖 | 分子量: 38998.078 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #47: RNA鎖 | 分子量: 24485.539 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #49: RNA鎖 | 分子量: 162190.062 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) HEPATITIS C VIRUS (ウイルス) |
| #50: RNA鎖 | 分子量: 602776.875 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
+60S RIBOSOMAL PROTEIN ... , 42種, 42分子 AAABACADAEAFAGAHAIAJALAMANAOAPAQARASATAUAVAWAXAYAZAaAbAcAdAe...
-タンパク質 , 2種, 2分子 AmBB
| #41: タンパク質 | 分子量: 14758.394 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #48: タンパク質 | 分子量: 70788.000 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
+40S RIBOSOMAL PROTEIN ... , 33種, 33分子 CACBCCCDCECFCGCHCICJCKCLCMCNCOCPCQCRCSCTCUCVCWCXCYCZCaCbCcCd...
-非ポリマー , 2種, 2分子 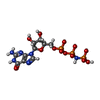

| #84: 化合物 | ChemComp-GNP /  分子量: 522.196 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H17N6O13P3 分子量: 522.196 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H17N6O13P3コメント: GppNHp, GMPPNP, エネルギー貯蔵分子類似体*YM |
|---|---|
| #85: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg |
-詳細
| Has protein modification | Y |
|---|---|
| 配列の詳細 | CHAIN 2 1-5025 GB M11167 1-5025 CHAIN 3 1-181 GB CR749662 1-181 CHAIN 4 1-121 GB X51545 274-394 |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: HCV-IRES 80S INITIATION COMPLEX WITH EIF5B PRE- LIKE STATE タイプ: RIBOSOME |
|---|---|
| 緩衝液 | 名称: 20MM TRIS-HCL, 5MM MGCL2, 100MM KCL, 0.2MM SPERMIDINE, 2MM DTT pH: 7.6 詳細: 20MM TRIS-HCL, 5MM MGCL2, 100MM KCL, 0.2MM SPERMIDINE, 2MM DTT |
| 試料 | 濃度: 0.15 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | 詳細: HOLEY CARBON |
| 急速凍結 | 装置: FEI VITROBOT MARK II / 凍結剤: ETHANE 詳細: VITRIFICATION 1 -- CRYOGEN- ETHANE, HUMIDITY- 100, TEMPERATURE- 93, INSTRUMENT- FEI VITROBOT MARK II, METHOD- BLOT FOR 2-4 SECONDS BEFORE PLUNGING, |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Tecnai F30 / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TECNAI F30 / 日付: 2012年12月22日 / 詳細: AUTOMATED DATA COLLECTION USING LEGINON |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 115000 X / 倍率(補正後): 194805 X / 最大 デフォーカス(公称値): 4500 nm / 最小 デフォーカス(公称値): 2000 nm / Cs: 2 mm |
| 撮影 | 電子線照射量: 20 e/Å2 フィルム・検出器のモデル: TVIPS TEMCAM-F416 (4k x 4k) |
| 放射波長 | 相対比: 1 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | 詳細: CTFFIND3 | ||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||
| 3次元再構成 | 解像度: 8.9 Å / 粒子像の数: 541570 / ピクセルサイズ(公称値): 2.37 Å / ピクセルサイズ(実測値): 2.37 Å 詳細: SUBMISSION BASED ON EXPERIMENTAL DATA FROM EMDB EMD-2682. (DEPOSITION ID: 12562). 対称性のタイプ: POINT | ||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL / 詳細: METHOD--RIGID BODY | ||||||||||||
| 原子モデル構築 | PDB-ID: 4CXC 4cxc Accession code: 4CXC / Source name: PDB / タイプ: experimental model | ||||||||||||
| 精密化 | 最高解像度: 8.9 Å | ||||||||||||
| 精密化ステップ | サイクル: LAST / 最高解像度: 8.9 Å
|
