BIM-BCL-xl complex / BIM-BCL-2 complex / regulation of developmental pigmentation / RUNX3 regulates BCL2L11 (BIM) transcription / positive regulation of mitochondrial membrane permeability involved in apoptotic process / apoptotic process involved in embryonic digit morphogenesis / positive regulation of fibroblast apoptotic process / positive regulation of endoplasmic reticulum stress-induced intrinsic apoptotic signaling pathway / Activation of BIM and translocation to mitochondria / developmental pigmentation ...BIM-BCL-xl complex / BIM-BCL-2 complex / regulation of developmental pigmentation / RUNX3 regulates BCL2L11 (BIM) transcription / positive regulation of mitochondrial membrane permeability involved in apoptotic process / apoptotic process involved in embryonic digit morphogenesis / positive regulation of fibroblast apoptotic process / positive regulation of endoplasmic reticulum stress-induced intrinsic apoptotic signaling pathway / Activation of BIM and translocation to mitochondria / developmental pigmentation / ear development / symbiont-mediated suppression of host apoptosis / BH3-only proteins associate with and inactivate anti-apoptotic BCL-2 members / regulation of organ growth / positive regulation of T cell apoptotic process / tube formation / mammary gland development / meiosis I / myeloid cell homeostasis / Bcl-2 family protein complex / cellular response to glucocorticoid stimulus / odontogenesis of dentin-containing tooth / NRAGE signals death through JNK / thymocyte apoptotic process / Deregulated CDK5 triggers multiple neurodegenerative pathways in Alzheimer's disease models / FOXO-mediated transcription of cell death genes / B cell homeostasis / positive regulation of IRE1-mediated unfolded protein response / T cell homeostasis / positive regulation of release of cytochrome c from mitochondria / host cell mitochondrion / host cell membrane / spleen development / extrinsic apoptotic signaling pathway in absence of ligand / positive regulation of intrinsic apoptotic signaling pathway / endomembrane system / FLT3 Signaling / positive regulation of cell cycle / thymus development / post-embryonic development / response to endoplasmic reticulum stress / cell-matrix adhesion / kidney development / positive regulation of protein-containing complex assembly / male gonad development / intrinsic apoptotic signaling pathway in response to DNA damage / Signaling by BRAF and RAF1 fusions / positive regulation of neuron apoptotic process / in utero embryonic development / regulation of apoptotic process / spermatogenesis / microtubule binding / mitochondrial outer membrane / positive regulation of apoptotic process / apoptotic process / protein kinase binding / mitochondrion / cytosol Similarity search - Function
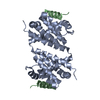
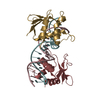
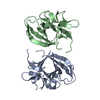
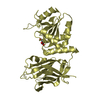
Journal: Acta Crystallogr D Biol Crystallogr / Year: 2015 Title: Structural basis of Deerpox virus-mediated inhibition of apoptosis. Authors: Denis R Burton / Sofia Caria / Bevan Marshall / Michele Barry / Marc Kvansakul / Abstract: Apoptosis is a key innate defence mechanism to eliminate virally infected cells. To counteract premature host-cell apoptosis, poxviruses have evolved numerous molecular strategies, including the use ...Apoptosis is a key innate defence mechanism to eliminate virally infected cells. To counteract premature host-cell apoptosis, poxviruses have evolved numerous molecular strategies, including the use of Bcl-2 proteins, to ensure their own survival. Here, it is reported that the Deerpox virus inhibitor of apoptosis, DPV022, only engages a highly restricted set of death-inducing Bcl-2 proteins, including Bim, Bax and Bak, with modest affinities. Structural analysis reveals that DPV022 adopts a Bcl-2 fold with a dimeric domain-swapped topology and binds pro-death Bcl-2 proteins via two conserved ligand-binding grooves found on opposite sides of the dimer. Structures of DPV022 bound to Bim, Bak and Bax BH3 domains reveal that a partial obstruction of the binding groove is likely to be responsible for the modest affinities of DPV022 for BH3 domains. These findings reveal that domain-swapped dimeric Bcl-2 folds are not unusual and may be found more widely in viruses. Furthermore, the modest affinities of DPV022 for pro-death Bcl-2 proteins suggest that two distinct classes of anti-apoptotic viral Bcl-2 proteins exist: those that are monomeric and tightly bind a range of death-inducing Bcl-2 proteins, and others such as DPV022 that are dimeric and only bind a very limited number of death-inducing Bcl-2 proteins with modest affinities.
Type: ADSC QUANTUM 315r / Detector: CCD / Date: Mar 1, 2013 / Details: SILICON MIRRORS (ADAPTIVE AND U-BENT)
Radiation
Monochromator: DOUBLE CRYSTAL MONOCHROMATOR (SI111) / Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray
Radiation wavelength
ID
Wavelength (Å)
Relative weight
1
0.9537
1
2
1.5498
1
Reflection
Resolution: 2.7→94.39 Å / Num. obs: 6039 / % possible obs: 100 % / Observed criterion σ(I): 2 / Redundancy: 10 % / Biso Wilson estimate: 65.25 Å2 / Rmerge(I) obs: 0.08 / Net I/σ(I): 19.5
Reflection shell
Resolution: 2.7→2.83 Å / Redundancy: 10.5 % / Rmerge(I) obs: 0.89 / Mean I/σ(I) obs: 2.8 / % possible all: 100
-
Processing
Software
Name
Version
Classification
PHENIX
(PHENIX.REFINE)
refinement
XDS
datareduction
Aimless
datascaling
SHELX
D
phasing
Refinement
Method to determine structure: SIRAS Starting model: NONE Resolution: 2.7→19.99 Å / SU ML: 0.39 / σ(F): 2 / Phase error: 29.54 / Stereochemistry target values: ML Details: RESIDUES FROM 1 AND 2 AND FROM 138 TO 155 ARE MISSING IN DPV022. IN BIM,THE RESIDUES 50 AND 55 AS WELL AS 74-76
Rfactor
Num. reflection
% reflection
Rfree
0.2206
273
4.6 %
Rwork
0.2051
-
-
obs
0.2059
5980
99.85 %
Solvent computation
Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL
Displacement parameters
Biso mean: 91.4 Å2
Refinement step
Cycle: LAST / Resolution: 2.7→19.99 Å
Protein
Nucleic acid
Ligand
Solvent
Total
Num. atoms
1271
0
0
7
1278
Refine LS restraints
Refine-ID
Type
Dev ideal
Number
X-RAY DIFFRACTION
f_bond_d
0.003
1288
X-RAY DIFFRACTION
f_angle_d
0.567
1731
X-RAY DIFFRACTION
f_dihedral_angle_d
14.347
493
X-RAY DIFFRACTION
f_chiral_restr
0.023
197
X-RAY DIFFRACTION
f_plane_restr
0.002
217
LS refinement shell
Resolution (Å)
Rfactor Rfree
Num. reflection Rfree
Rfactor Rwork
Num. reflection Rwork
Refine-ID
% reflection obs (%)
2.7004-3.3995
0.3499
128
0.2532
2780
X-RAY DIFFRACTION
100
3.3995-19.9904
0.1938
145
0.1905
2927
X-RAY DIFFRACTION
100
Refinement TLS params.
Method: refined / Refine-ID: X-RAY DIFFRACTION
ID
L11 (°2)
L12 (°2)
L13 (°2)
L22 (°2)
L23 (°2)
L33 (°2)
S11 (Å °)
S12 (Å °)
S13 (Å °)
S21 (Å °)
S22 (Å °)
S23 (Å °)
S31 (Å °)
S32 (Å °)
S33 (Å °)
T11 (Å2)
T12 (Å2)
T13 (Å2)
T22 (Å2)
T23 (Å2)
T33 (Å2)
Origin x (Å)
Origin y (Å)
Origin z (Å)
1
2.1942
0.6479
-0.1852
5.2644
3.2183
2.9959
-0.3855
0.1817
0.1259
-0.5382
0.2095
0.4209
0.0537
0.1091
0.3587
0.6688
0.0442
-0.1142
0.8037
-0.0503
0.398
74.499
13.2418
19.8123
2
6.2506
2.7903
1.5516
6.8214
2.0327
7.4543
0.1397
-0.2278
0.882
0.443
0.1958
-0.153
-0.6046
0.9269
-0.3767
0.6635
-0.1318
0.0505
0.7538
-0.0884
0.5003
89.0535
27.8387
4.7412
3
6.3632
1.8678
-1.6277
5.1342
-4.4769
3.804
0.0966
-1.0878
1.0907
0.7883
0.3098
-0.0666
-0.3968
0.115
-0.462
0.6841
-0.0787
0.0351
0.5481
-0.209
0.5819
84.5326
27.3331
9.3474
4
3.4885
-2.6896
1.6542
4.1673
0.712
2.9242
0.0179
1.2571
1.1387
0.0067
-0.6907
-0.1777
-0.0553
-0.6333
0.6187
1.0567
-0.0426
0.1401
0.7286
0.0255
0.6499
84.4302
25.0992
-6.5493
5
2.7041
-0.1705
1.2253
3.3683
-3.6906
4.4318
-0.5951
1.5805
1.8084
-0.4363
0.6493
-0.2362
-1.6467
2.4872
0.3693
1.026
-0.1709
-0.1579
1.4591
-0.0688
1.2381
97.177
32.8979
-0.5536
Refinement TLS group
ID
Refine-ID
Refine TLS-ID
Selection details
1
X-RAY DIFFRACTION
1
CHAIN 'A' AND (RESID3THROUGH30 )
2
X-RAY DIFFRACTION
2
CHAIN 'A' AND (RESID31THROUGH87 )
3
X-RAY DIFFRACTION
3
CHAIN 'A' AND (RESID88THROUGH117 )
4
X-RAY DIFFRACTION
4
CHAIN 'A' AND (RESID118THROUGH138 )
5
X-RAY DIFFRACTION
5
CHAIN 'B' AND (RESID56THROUGH73 )
+
About Yorodumi
-
News
-
Feb 9, 2022. New format data for meta-information of EMDB entries
New format data for meta-information of EMDB entries
Version 3 of the EMDB header file is now the official format.
The previous official version 1.9 will be removed from the archive.
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information Homo sapiens (human)
Homo sapiens (human) X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Citation
Citation

 Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads


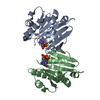
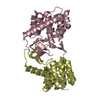
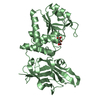


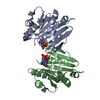
 PDBj
PDBj








 Assembly
Assembly

 Deerpox virus (strain W-1170-84) / Strain: W-1170-84 / Gene: DpV84gp022 / Plasmid: pDUET / Production host:
Deerpox virus (strain W-1170-84) / Strain: W-1170-84 / Gene: DpV84gp022 / Plasmid: pDUET / Production host: 
 Mass: 18.015 Da / Num. of mol.: 7 / Source method: isolated from a natural source / Formula: H2O
Mass: 18.015 Da / Num. of mol.: 7 / Source method: isolated from a natural source / Formula: H2O Sample preparation
Sample preparation Processing
Processing