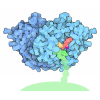
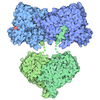
- PDB-4mko: Crystal structure of the monomeric, cleaved form of the Pore-Form... -
+
データを開く
IDまたはキーワード:
読み込み中...
-
基本情報
登録情報
データベース: PDB / ID: 4mko
タイトル
Crystal structure of the monomeric, cleaved form of the Pore-Forming Toxin Monalysin
要素
Monalysin
キーワード
TOXIN / Pore-Forming Toxin
機能・相同性
機能・相同性情報
hemolysis in another organism / porin activity / pore complex / monoatomic ion transport / protein homooligomerization / toxin activity / host cell plasma membrane / extracellular region 類似検索 - 分子機能
ジャーナル: J Biol Chem / 年: 2015 タイトル: X-ray and Cryo-electron Microscopy Structures of Monalysin Pore-forming Toxin Reveal Multimerization of the Pro-form. 著者: Philippe Leone / Cecilia Bebeacua / Onya Opota / Christine Kellenberger / Bruno Klaholz / Igor Orlov / Christian Cambillau / Bruno Lemaitre / Alain Roussel / 要旨: β-Barrel pore-forming toxins (β-PFT), a large family of bacterial toxins, are generally secreted as water-soluble monomers and can form oligomeric pores in membranes following proteolytic cleavage ...β-Barrel pore-forming toxins (β-PFT), a large family of bacterial toxins, are generally secreted as water-soluble monomers and can form oligomeric pores in membranes following proteolytic cleavage and interaction with cell surface receptors. Monalysin has been recently identified as a β-PFT that contributes to the virulence of Pseudomonas entomophila against Drosophila. It is secreted as a pro-protein that becomes active upon cleavage. Here we report the crystal and cryo-electron microscopy structure of the pro-form of Monalysin as well as the crystal structures of the cleaved form and of an inactive mutant lacking the membrane-spanning region. The overall structure of Monalysin displays an elongated shape, which resembles those of β-pore-forming toxins, such as Aerolysin, but is devoid of a receptor-binding domain. X-ray crystallography, cryo-electron microscopy, and light-scattering studies show that pro-Monalysin forms a stable doughnut-like 18-mer complex composed of two disk-shaped nonamers held together by N-terminal swapping of the pro-peptides. This observation is in contrast with the monomeric pro-form of the other β-PFTs that are receptor-dependent for membrane interaction. The membrane-spanning region of pro-Monalysin is fully buried in the center of the doughnut, suggesting that upon cleavage of pro-peptides, the two disk-shaped nonamers can, and have to, dissociate to leave the transmembrane segments free to deploy and lead to pore formation. In contrast with other toxins, the delivery of 18 subunits at once, nearby the cell surface, may be used to bypass the requirement of receptor-dependent concentration to reach the threshold for oligomerization into the pore-forming complex.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Pseudomonas entomophila (バクテリア)
Pseudomonas entomophila (バクテリア) X線回折 /
X線回折 /  データ登録者
データ登録者 引用
引用

 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード








 PDBj
PDBj









 集合体
集合体





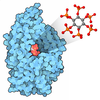
 分子量: 200.590 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Hg
分子量: 200.590 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Hg
 分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn
分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn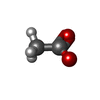
 分子量: 59.044 Da / 分子数: 13 / 由来タイプ: 合成 / 式: C2H3O2
分子量: 59.044 Da / 分子数: 13 / 由来タイプ: 合成 / 式: C2H3O2 分子量: 18.015 Da / 分子数: 1318 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 1318 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 解析
解析